1.8. яРЮПЕМХЕ ЙКЕРНЙ in vitro
йКЕРЙХ ЯРЮПЕЧР МЕ РНКЭЙН in vivo, МН Х in vitro. аНКЕЕ РНЦН, Б СЯКНБХЪУ in vitro НЯНАЕММН НРВ╦РКХБН ОПНЪБКЪЕРЯЪ ПНКЭ ЦХОЕПНЙЯХХ √ ЕЯРЕЯРБЕММНЦН Х, ОН-БХДХЛНЛС, ЕДХМЯРБЕММНЦН Б ЩРХУ СЯКНБХЪУ ТЮЙРНПЮ ХУ ЯРЮПЕМХЪ.
1.8.1.
йЮЙ ХГБЕЯРМН, БШПЮЫХБЮМХЕ ЙКЕРНЙ БМЕ НПЦЮМХГЛЮ ОПНБНДХРЯЪ Б ЯОЕЖХЮКЭМШУ ЯНЯСДЮУ (ТКЮЙНМЮУ) ОПХ ЮРЛНЯТЕПМНЛ ДЮБКЕМХХ Х, ЯКЕДНБЮРЕКЭМН, ОПХ Пн2, ГМЮВХРЕКЭМН ОПЕБНЯУНДЪЫЕЛ ГМЮВЕМХЪ, ЙНРНПШЕ СЯРЮМЮБКХБЮЧРЯЪ Б НПЦЮМХГЛЕ Б МНПЛЕ. нАШВМН Б ХМЙСАЮЖХНММНИ ФХДЙНЯРХ Пн2 АКХГЙН Й Пн2 БНГДСУЮ. лНКЕЙСКШ н2 ВЕПЕГ РНМЙХИ ЯКНИ ОХРЮРЕКЭМНИ ЯПЕДШ БН ТКЮЙНМЕ ЯБНАНДМН ДХТТСМДХПСЧР Й ЙКЕРЙЮЛ Х БМСРПХ МХУ СЯРЮМЮБКХБЮЕРЯЪ БШЯНЙНЕ Пн2, МЕБНГЛНФМНЕ in vivo ХКХ, БН БЯЪЙНЛ ЯКСВЮЕ, ОПЕБШЬЮЧЫЕЕ ДНОСЯРХЛШЕ ГМЮВЕМХЪ.я РНВЙХ ГПЕМХЪ ЙХЯКНПНДМН-ОЕПЕЙХЯМНИ ЙНМЖЕОЖХХ ЯРЮПЕМХЪ, СЯКНБХЪ in vitro ОПЕДЯРЮБКЪЧРЯЪ АНКЕЕ ВЕЛ ОНДУНДЪЫХЛХ ДКЪ ХГСВЕМХЪ ОПНЖЕЯЯЮ ЯРЮПЕМХЪ ЙКЕРНЙ, РЮЙ ЙЮЙ Б ЩРХУ СЯКНБХЪУ НМ ОПНРЕЙЮЕР ХМРЕМЯХБМЕЕ, Б СЯЙНПЕММНЛ РЕЛОЕ Х, ВРН НВЕМЭ БЮФМН, Б ╚ВХЯРНЛ╩ БХДЕ, Р.Е. ОПХ ОНКМНЛ НРЯСРЯРБХХ ЙЮЙНЦН-КХАН БКХЪМХЪ ЯХЯРЕЛ НПЦЮМХГЛЮ, ЙНРНПНЕ ХЛЕЕР ЛЕЯРН ОПХ ЯРЮПЕМХХ in vivo. дЮММНЕ НАЯРНЪРЕКЭЯРБН ЯПЮГС ЯРЮБХР ЛМНЦХЕ РЕНПХХ ЯРЮПЕМХЪ Б ПЮГПЪД БРНПНЯРЕОЕММШУ ХКХ ЯСЦСАН СЛНГПХРЕКЭМШУ, ОНЯЙНКЭЙС БНГПЮЯРМШЕ ХГЛЕМЕМХЪ ОПНХЯУНДЪР ХКХ ЛНЦСР ОПНХЯУНДХРЭ Х АЕГ ПЕЮКХГЮЖХХ ОНЯРСКХПСЕЛШУ Б МХУ ОНКНФЕМХИ. оПХДЮМХЕ МЮЛХ ЯРНКЭ БЮФМНЦН ГМЮВЕМХЪ ТЕМНЛЕМС ЯРЮПЕМХЪ ЙКЕРНЙ in vitro ЯБЪГЮМН Я РЕЛ, ВРН ХЛЕММН Б ЩРХУ ╚ОПНЯРШУ╩ СЯКНБХЪУ ЛНФМН АСДЕР ЯЙНПЕЕ Х Я ЛЕМЭЬХЛХ РПСДМНЯРЪЛХ ОНГМЮРЭ ТХГХЙН-УХЛХВЕЯЙХЕ НЯМНБШ ЯРЮПЕМХЪ Х ЯСЫМНЯРЭ АХНКНЦХХ ЩРНЦН ОПНЖЕЯЯЮ БННАЫЕ.
б МЮЯРНЪЫЕЕ БПЕЛЪ, НДМЮЙН, МЕ ЯСЫЕЯРБСЕР ЕДХМНЦН ЛМЕМХЪ ОН ОНБНДС НАЫМНЯРХ ОПХВХМ Х ЛЕУЮМХГЛНБ ЯРЮПЕМХЪ ЙКЕРНВМШУ ЙСКЭРСП Х ЯРЮПЕМХЪ ЙКЕРНЙ Б ЯНЯРЮБЕ ЛМНЦНЙКЕРНВМНЦН НПЦЮМХГЛЮ, Н ВЕЛ ЯБХДЕРЕКЭЯРБСЧР ОПНРХБНОНКНФМШЕ РНВЙХ ГПЕМХЪ Б КХРЕПЮРСПЕ (йЮОХРЮМНБ, 1986). йЮМСМЦН (Kanungo, 1982), МЮОПХЛЕП, УНРЪ Х ЯВХРЮЕР, ВРН ОПХВХМЮ ЯРЮПЕМХЪ НПЦЮМХГЛЮ ЯНЯРНХР Б ЯРЮПЕМХХ ЕЦН ЙКЕРНЙ, БЛЕЯРЕ Я РЕЛ ОНКЮЦЮЕР: ╚СЯКНБХЪ in vitro МЕ ЯННРБЕРЯРБСЧР ТХГХНКНЦХВЕЯЙХЛ Х ЯБНИЯРБЮ ЙКЕРНЙ ЛНЦСР НЙЮГЮРЭЯЪ ХГЛЕМЕММШЛХ.
еЯКХ ХЯЯКЕДНБЮМХЪ in vitro Х ДЮЧР МЕЙНРНПСЧ ОНКЕГМСЧ ХМТНПЛЮЖХЧ Н ЯЮЛНИ ЙКЕРЙЕ, РН НМХ ХЛЕЧР НЦПЮМХВЕММСЧ ЖЕММНЯРЭ, ЙНЦДЮ ПЕВЭ ХД╦Р Н ЯРЮПЕМХХ НПЦЮМХГЛЮ Б ЖЕКНЛ╩. я ОПХБЕДЕММШЛ БШЯЙЮГШБЮМХЕЛ ЛНФМН ЯНЦКЮЯХРЭЯЪ КХЬЭ НРВЮЯРХ. дЕИЯРБХРЕКЭМН, ЯРЮПЕМХЕ ЙКЕРНЙ in vitro МЕ ЛНФЕР НРПЮГХРЭ БЕЯЭ ЯКНФМШИ ЯОЕЙРП БНГПЮЯРМШУ ХГЛЕМЕМХИ, ОПНХЯУНДЪЫХУ Б ЖЕКНЯРМНЛ НПЦЮМХГЛЕ МЮ БЯЕУ СПНБМЪУ Х, Й РНЛС ФЕ, Б МЕЛЮКНИ ЯРЕОЕМХ НОПЕДЕКЪЕЛШУ ЯХЯРЕЛНИ ПЮГКХВМШУ ЯБЪГЕИ Б МЕЛ, Б РНЛ ВХЯКЕ НАПЮРМШУ. оПХЛЕМХРЕКЭМН Й СЯКНБХЪЛ in vitro РЕПЪЕР ЯЛШЯК ПЪД ОПХМЖХОНБ ЯРЮПЕМХЪ, ОПНЪБКЪЧЫХУЯЪ МЮ НПЦЮМХГЛЕММНЛ СПНБМЕ (ЯЛ. О. 1.1.2), МН МЕЙНРНПШЕ ХГ МХУ ОПНДНКФЮЧР ДЕИЯРБНБЮРЭ Х Б ЙКЕРНВМШУ ЙСКЭРСПЮУ. рЮЙНБШ, Б ВЮЯРМНЯРХ, ЛМНЦННВЮЦНБНЯРЭ ОПНЖЕЯЯЮ ЯРЮПЕМХЪ, Р.Е. ПЮГБХРХЕ ОНБПЕФДЕМХИ Б ПЮГКХВМШУ ВЮЯРЪУ ЙКЕРЙХ ХКХ Б ПЮГКХВМШУ ЕЕ ЛНКЕЙСКЪПМШУ ЖХЙКЮУ, Х ЦЕРЕПНУПНММНЯРЭ ЯРЮПЕМХЪ ЯПЕДХ ЙКЕРНЙ НДМНЦН ЙСКЭРХБХПСЕЛНЦН РХОЮ. йПНЛЕ РНЦН, Б ЩРХУ СЯКНБХЪУ ОПХМЖХОШ МЕНАПЮРХЛНЯРХ, МЕПЕЦСКХПСЕЛНЯРХ Х МЕОПЕПШБМНЯРХ ЯРЮПЕМХЪ ЙКЕРНЙ ДНКФМШ, НВЕБХДМН, ОПНЪБКЪРЭЯЪ АНКЕЕ ЪБЯРБЕММН.сЙЮГЮММШЕ БШЬЕ МЕДНЯРЮРЙХ ОПХ ХГСВЕМХХ ЯРЮПЕМХЪ ЙКЕРНЙ БМЕ НПЦЮМХГЛЮ МЕ ОПЕДЯРЮБКЪЧРЯЪ ОПХМЖХОХЮКЭМШЛХ, ЕЯКХ ХЛЕРЭ Б БХДС, ВРН НДМЮ ХГ НЯМНБМШУ ГЮДЮВ ЦЕПНМРНКНЦХХ ЯНЯРНХР Б СЯРЮМНБКЕМХХ ЦКЮБМНЦН ОЕПБХВМНЦН ТЮЙРНПЮ НЙ-ПСФЮЧЫЕИ ЯПЕДШ, ОПЕДНОПЕДЕКЪЧЫЕЦН ЯРЮПЕМХЕ БЯЕУ ФХБШУ НПЦЮМХГЛНБ. рЮЙХЛ ТЮЙРНПНЛ, ЙЮЙ ЛШ ОНКЮЦЮЕЛ, ЪБКЪЕРЯЪ ЦХОЕПНЙЯХЪ Б ГЕЛМНИ ЮРЛНЯТЕПЕ, ОНЩРНЛС ФХГМЭ ЙКЕРНЙ Б СЯКНБХЪУ in vitro ЛНФМН ЯВХРЮРЭ СДНАМНИ ЩЙЯОЕПХЛЕМРЮКЭМНИ ЛНДЕКЭЧ ДКЪ ХГСВЕМХЪ ДЕИЯРБХЪ ХЛЕММН СЙЮГЮММНЦН ТХГХВЕЯЙНЦН ТЮЙРНПЮ МЮ ХУ ЯРЮПЕМХЕ. нАШВМНЕ 18-21 %-МНЕ ЯНДЕПФЮМХЕ н2 Б БНГДСУЕ Х ЯННРБЕРЯРБЕММН БШЯНЙХЕ СПНБМХ ДХЯАЮКЮМЯЮ Δ (он √ юн) Х ОЕПНЙЯХЦЕМЮГМШУ ОПНЖЕЯЯНБ НЙЮГШБЮЧР СЦМЕРЮЧЫЕЕ ДЕИЯРБХЕ МЮ ЯСАЙКЕРНВМШЕ ЩКЕЛЕМРШ, МЮ МНПЛЮКЭМШЕ ТХГХНКНЦХВЕЯЙХЕ Х ЛЕРЮАНКХВЕЯЙХЕ ОПНЖЕЯЯШ. б ПЕГСКЭРЮРЕ ОНЯКЕДМХЕ ОНЯРЕОЕММН ГЮРСУЮЧР, Х АНКЭЬХМЯРБН ЙКЕРНЙ ОНЦХАЮЕР БЯКЕДЯРБХЕ НЙХЯКХРЕКЭМНЦН ЖХРНКХГЮ ХКХ ОН ЙХЯКНПНДМН-ОЕПЕЙХЯМНЛС ЛЕУЮМХГЛС ЮОНОРНГЮ (ЯЛ. О. 7.1).
тЮЙРНБ, СЙЮГШБЮЧЫХУ МЮ БЕДСЫСЧ ПНКЭ ХГАШРНВМШУ Пн2, ютй Х онк Б ЯМХФЕМХХ БШФХБЮЕЛНЯРХ ЙКЕРНЙ Б СЯКНБХЪУ in vitro Х ОПНРЕЙРНПМНЦН ДЕИЯРБХЪ ПЮГКХВМШУ ЮМРХНЙЯХДЮМРМШУ ТЮЙРНПНБ, АНКЕЕ, ВЕЛ ДНЯРЮРНВМН (Branton et al., 1998; Drukarch et al., 1998; Heppner et al., 1998).
й ВХЯКС ОНЯКЕДМХУ НРМЕЯЕМ МЕДЮБМН Х L-ЙЮПМНГХМ. дНАЮБКЕМХЕ ТХГХНКНЦХВЕЯЙХУ ЙНМЖЕМРПЮЖХИ ЕЦН Й ЯРЮМДЮПРМШЛ ЯПЕДЮЛ СБЕКХВХБЮЕР ОПНДНКФХРЕКЭМНЯРЭ ФХГМХ ТХАПНАКЮЯРНБ ВЕКНБЕ-ЙЮ in vitro Х ГЮЛЕДКЪЕР ОПНЖЕЯЯШ ТХГХНКНЦХВЕЯЙНЦН ЯРЮПЕМХЪ Б МХУ. дКХРЕКЭМН ОЮЯЯХПСЕЛШЕ МЮ НАШВМШУ ЯПЕДЮУ ЙКЕРЙХ ОНЯКЕ ОЕПЕМНЯЮ ХУ Б ЙЮПМНГХМ-ЯНДЕП-ФЮЫСЧ ЯПЕДС ОПНЪБКЪКХ НЛНКЮФХБЮЧЫХИ ЩТТЕЙР. нОРХВЕЯЙХИ ФЕ ХГНЛЕП D-ЙЮПМНГХМ МЕ НАКЮДЮК СЙЮГЮММШЛХ ЯБНИЯРБЮЛХ (уНККХДЕИ, лЮЙтЮПКЮМД, 2000) б РН ФЕ БПЕЛЪ Б УНДЕ ДКХРЕКЭМНЦН ЙСКЭРХБХПНБЮМХЪ НОПЕДЕК╦ММШИ ОПНЖЕМР ЙКЕРНЙ МЕ РНКЭЙН МЕ ДЕЦПЮДХПСЕР, МН Х, ЮДЮОРХПСЪЯЭ Й РНЙЯХВЕЯЙХЛ НЙХЯКХРЕКЭМШЛ СЯКНБХЪЛ, ╚ДНАХБЮЕРЯЪ╩ РНЦН, ВРН БМСРПХЙКЕРНВМШИ ОЮПЮЛЕРП Δ (он √ юн) МЕ ОНБШЬЮЕРЯЪ ДН БШЯНЙХУ ГМЮВЕМХИ Δю2 ХКХ Δж, МН ЛНФЕР НЯРЮМНБХРЭЯЪ МЮ МЕЯЙНКЭЙН ЛЕМЭЬЕЛ СПНБМЕ Δй, МЕНАУНДХЛНЛ ДКЪ ХУ ГКНЙЮВЕЯРБЕММНИ РПЮМЯТНПЛЮЖХХ. яКСВЮХ ╚ЯОНМРЮММНИ╩ ЛЮКХЦМХГЮЖХХ ЙКЕРНЙ Б ЙСКЭРСПЕ Х БНГЛНФМШИ ЕЕ ЛЕУЮМХГЛ НАЯСФДЮЧРЯЪ МЮЛХ НРДЕКЭМН Б ЦКЮБЕ 4.1.8.2. оПХБЕДЕММШЕ БШЬЕ ЯННАПЮФЕМХЪ ЛНФМН ЯВХРЮРЭ ВЮЯРЭЧ МЮЬХУ РЕНПЕРХВЕЯЙХУ ОНКНФЕМХИ Н ОПХВХМЮУ Х ЯКЕДЯРБХЪУ ЯРЮПЕМХЪ ЙКЕРНЙ in vitro. дКЪ ОНДРБЕПФДЕМХЪ Х ПЮГБХРХЪ ЩРХУ ОНКНФЕМХИ ЕЯРЕЯРБЕММН ОПХБКЕВЭ МЕЙНРНПШЕ СФЕ ХГБЕЯРМШЕ ТЮЙРШ, ЯНДЕПФЮМХЕ Х ЯЛШЯК ЙНРНПШУ КЕЦЙН ЛНЦСР АШРЭ ╚БОХЯЮМШ╩ Б ЙХЯКНПНДМН-ОЕПЕЙХЯМСЧ ЙНМЖЕОЖХЧ ЯРЮПЕМХЪ ЙКЕРНЙ. мЮВМЕЛ Я РНЦН, ВРН БШЬЕНОХЯЮММШЕ НАШВМШЕ СЯКНБХЪ ЙСКЭРХБХПНБЮМХЪ ЙКЕРНЙ, ЪБКЪЧЫХЕЯЪ ДКЪ МХУ РНЙЯХВЕЯЙХЛХ, ЛНЦСР АШРЭ ЯЛЪЦВЕМШ ОСРЕЛ ХЯЙСЯЯРБЕММНЦН ЯМХФЕМХЪ ЙНМЖЕМРПЮЖХХ н2 Б ЦЮГНБНИ ЯПЕДЕ. оПХ ЩРНЛ СЦМЕРЮЧЫЕЕ ДЕИЯРБХЕ ЦХОЕПНЙЯХХ Х ЯЙНПНЯРЭ ЯРЮПЕМХЪ ЙКЕРНЙ ДНКФМШ ЯМХГХРЭЯЪ. яКЕДСЕР РЮЙФЕ ХЛЕРЭ Б БХДС, ВРН РЮЙЮЪ ХГБЕЯРМЮЪ АХНКНЦХВЕЯЙЮЪ ЙНМЯРЮМРЮ, ЙЮЙ КХЛХР уЕИТКХЙЮ, Б ДЕИЯРБХРЕКЭМНЯРХ НЙЮГЮКЮЯЭ ОЕПЕЛЕММНИ БЕКХВХМНИ, ГЮБХЯЪЫЕИ НР ЯНДЕПФЮМХЪ н2 Б ЦЮГНБНИ ЯПЕДЕ, ОПХВЕЛ Б СЯКНБХЪУ НЙЯХДЮРХБМНЦН ЯРПЕЯЯЮ ЩРНР КХЛХР СЛЕМЭЬЮЕРЯЪ, Ю ОПХ ЯМХФЕМХХ Пн2, МЮОПНРХБ, БНГПЮЯРЮЕР (Chen et al., 1995).
дЕИЯРБХРЕКЭМН, ОПЕАШБЮМХЕ ЙСКЭРСПШ ТХАПНАКЮЯРНБ Б ЮРЛНЯТЕПЕ Я ОНМХФЕММШЛ ЯНДЕПФЮМХЕЛ н2 (10 %) СДКХМЪЕР ЯПНЙ ХУ ФХГМХ МЮ 20-30 %.
рН ФЕ ОПНХЯУНДХР Х Я ЙКЕРЙЮЛХ КЕЦЙХУ ВЕКНБЕЙЮ Х ЛШЬЕИ (Packer, Walton, 1977). оЕПХНД ОПНКХТЕПЮРХБМНИ ФХГМЕЯОНЯНАМНЯРХ ДХОКНХДМШУ ТХАПНАКЮЯРНБ IMR90 ВЕКНБЕЙЮ Я ПЮГКХВМШЛХ ХЯУНДМШЛХ СПНБМЪЛХ СДБНЕМХЪ ОНОСКЪЖХХ СБЕКХВХБЮ-ЕРЯЪ ОПХ ЯМХФЕМХХ ЯНДЕПФЮМХЪ н2 Б ЯПЕДЕ ДН 1,6 ХКХ 12 %. щРНР ОЕПХНД ОПХ 1 % н2 БНГПЮЯРЮЕР МЮ 22 %, Ю БНГБПЮЫЕМХЕ ЙСКЭРСП ХГ ЯПЕДШ Я 1 % н2 Б ЯПЕДС Я 20 % н2 АШЯРПН ПЮГБХБЮЕР ХУ ЯРЮПЕМХЕ. б ЙСКЭРСПЕ ДХОКНХДМШУ ТХАПНАКЮЯРНБ НР АНКЭМНЦН ЯХМДПНЛНЛ бЕПМЕПЮ (ПЮММЕЕ ЯРЮПЕМХЕ) ОПНДНКФХРЕКЭМНЯРЭ ПЕОКХЙЮРХБМНИ ФХГМЕЯОНЯНАМНЯРХ РЮЙФЕ БНГПЮЯРЮЕР ОПХ ЯМХФЕМХХ Пн2 (Saito et al., 1995). гЮЛЕДКЕМХЕ ЯРЮПЕМХЪ ЙСКЭРХБХПСЕЛШУ УНМДПНЖХРНБ ЙСПХМНЦН ЩЛАПХНМЮ ОНЙЮГЮМН ОПХ 8 %-МНЛ ЯНДЕПФЮМХХ н2 Б ЮРЛНЯТЕПЕ ОН ЯПЮБМЕМХЧ Я ЙНМРПНКЕЛ (18 %), ОПХВ╦Л НОШРМШЕ ЙКЕРЙХ ДНКЭЬЕ ЯНУПЮМЪКХ ОПХГМЮЙХ ╚ЛНКНДШУ╩, ХЛЕКХ АНКЕЕ БШЯНЙСЧ ЯЙНПНЯРЭ ОПНКХТЕПЮЖХХ (Nevo et al., 1988). оНД БКХЪМХЕЛ ПЮГКХВМШУ ЮМРХНЙЯХДЮМРНБ ЯЙНПНЯРЭ ОПНКХТЕПЮЖХХ ЙКЕРНВМШУ ЙСКЭРСП РЮЙФЕ СБЕ-КХВХБЮЕРЯЪ, Ю ЯРЮПЕМХЕ ХУ ГЮЛЕДКЪЕРЯЪ (Packer, Walton, 1977; нАСУНБЮ, 1986), ВРН ОНДРБЕПФДЮЕР СФЕ ЯЙЮГЮММНЕ БШЬЕ: ЪБМН ХГАШРНВМНЕ ДЕИЯРБХЕ НЙЯХДЮМРНБ ОНДЮБКЪЕР ОПНКХТЕПЮЖХЧ ЙКЕРНЙ Х НАСЯКНБКХБЮЕР СЯЙНПЕММНЕ ХУ ЯРЮПЕМХЕ.б ЩЙЯОЕПХЛЕМРЮУ Я ЙКЕРНВМШЛХ ЙСКЭРСПЮЛХ ЯПЮБМХРЕКЭМН КЕЦЙН ОПНБЕПХРЭ РЮЙФЕ ДЕИЯРБХЕ н2-ГЮБХЯХЛНЦН ЛЕУЮМХГЛЮ ПЕЦСКЪЖХХ ЙНКХВЕЯРБЮ ДШУЮРЕКЭМШУ ТЕПЛЕМРНБ (Murphy et al., 1984; Suzuki et al., 1998) Х ЛХРНУНМДПХИ (нГЕПМЧЙ, 1978). яНЦКЮЯМН ЩРНЛС ЛЕУЮМХГЛС, ОПХ ОКЮБМНЛ Х ЛЕДКЕММНЛ БНГПЮЯРЮМХХ СПН-БМЪ ЦХОЕПНЙЯХХ ЯНДЕПФЮМХЕ РЮЙХУ ТЕПЛЕМРНБ Х ВХЯКН ЛХРНУНМДПХИ ДНКФМШ ОНЯРЕОЕММН МЮПЮЯРЮРЭ, Ю ОПХ ЦХОНЙЯХХ, МЮНАНПНР, √ ОЮДЮРЭ. дЕИЯРБХРЕКЭМН, ОПХ БШПЮЫХБЮМХХ ЙСКЭРСПШ ТХАПНАКЮЯРНБ МЮ ЯПЕДЕ Я ОНМХФЕММШЛ ЯНДЕПФЮМХЕЛ н2 ЙНМЖЕМРПЮЖХЪ ЖХРНУПНЛНБ ГМЮВХРЕКЭМН ЯМХФЮЕРЯЪ (Pius, 1970). гДЕЯЭ, АЕГСЯКНБМН, ГЮДЕИЯРБНБЮМ НАЗЕЙРХБМШИ ОПНЖЕЯЯ ЮДЮОРЮЖХХ ДШУЮРЕКЭМНИ ЯХЯРЕЛШ Й БМСРПХЙКЕРНВМНЛС СПНБМЧ Пн2. нДМЮЙН Б ДЮММНЛ ТЕМНЛЕМЕ МЕ ЛЕМЭЬЕЕ ГМЮВЕМХЕ ХЛЕЕР ЯЙНПНЯРЭ ЮДЮОРЮЖХХ, НР ЙНРНПНИ АСДЕР ГЮБХЯЕРЭ Х ХМРЕМЯХБМНЯРЭ ЯРЮПЕМХЪ ЙСКЭРХБХПСЕЛШУ ЙКЕРНЙ. йЮФЕРЯЪ НВЕБХДМШЛ, ВРН Б ОПНЖЕЯЯЕ АХНКНЦХВЕЯЙНИ ЩБНКЧЖХХ ЛМНЦНЙКЕРНВМШИ НПЦЮМХГЛ ОПХЯОНЯЮАКХБЮКЯЪ Й ОНЯРЕОЕММНЛС МЮПЮЯРЮМХЧ Пн2 Б ГЕЛМНИ ЮРЛНЯТЕПЕ РЮЙФЕ ОНЯРЕОЕММН.
оПХ ЩРНЛ БМСРПХ ЙКЕРНЙ ЯЮЛШЛ ЩТТЕЙРХБМШЛ ЛНФМН ЯВХРЮРЭ ╚ЛХРНУНМДПХЮКЭМШИ╩ ЛЕУЮМХГЛ ЮДЮОРЮЖХХ: ЙНКХВЕЯРБН ТЕПЛЕМРНБ ДШУЮРЕКЭМНИ ЖЕОХ Х ЯЮЛХУ ЛХРНУНМДПХИ БЮПЭХПСЕРЯЪ ЯЮЛННПЦЮМХГСЧЫЕИЯЪ ЯХЯРЕЛНИ РЮЙ, ВРНАШ НМН НАЕЯОЕВХБЮКН ЖЕКНЯРМНЯРЭ Х НРМНЯХРЕКЭМН МНПЛЮКЭМНЕ ТСМЙЖХНМХПНБЮМХЕ ЙКЕРНЙ ОПХ ХГЛЕМЕМХЪУ БМСРПХЙКЕРНВМНЦН Пн2 Б НОПЕДЕК╦ММШУ ЩБНКЧЖХНММН ЮОПНАХПНБЮММШУ СФЕ ОПЕДЕКЮУ.яНБЯЕЛ ХМЮЪ ЯХРСЮЖХЪ ЯЙКЮДШБЮЕРЯЪ ОПХ АШЯРПНЛ ОЕПЕМЕЯЕМХХ ЙКЕРНЙ ХГ ФХБНЦН НПЦЮМХГЛЮ Б СЯКНБХЪ in vitro. пЕГЙХИ ОЕПЕБНД ХУ Б ЯНЯРНЪМХЕ ЦХОЕПНЙЯХХ ПЮБМНЯХКЕМ МЮМЕЯЕМХЧ ХЛ ГМЮВХРЕКЭМНЦН ЯЙЮВЙННАПЮГМНЦН БНГЛСЫЮЧЫЕЦН БНГ-ДЕИЯРБХЪ, Й ЙНРНПНЛС НМХ, БННАЫЕ ЦНБНПЪ, МЕ ОНДЦНРНБКЕМШ. йЮЙ ФЕ ПЕЮЦХПСЕР ОЕПБХВМЮЪ ЙКЕРНВМЮЪ ЙСКЭРСПЮ МЮ РЮЙНЕ БНГЛСЫЕМХЕ? оН-БХДХЛНЛС, МЮ ОПНРЪФЕМХХ НОПЕДЕКЕММНЦН МЮВЮКЭМНЦН ОЕПХНДЮ ЙСКЭРСПЮКЭМЮЪ ЯПЕДЮ ЪБКЪЕРЯЪ ДКЪ ЙКЕРНЙ ╚ЯРПЕЯЯНБНИ╩, Ю ЯНЯРНЪМХЕ ЯЮЛХУ ЙКЕРНЙ Б ЩРНР ОЕПХНД √ ЬНЙНБШЛ. гЮРЕЛ ЙЮЙНЕ-РН БПЕЛЪ ГЮРПЮВХБЮЕРЯЪ МЮ ОНДЦНРНБЙС Х ОПНБЕДЕМХЕ БНГЛНФМШУ Б ЩРХУ ЩЙЯРПЕЛЮКЭМШУ СЯКНБХЪУ ЮДЮОРХБМШУ ╚ЛЕПНОПХЪРХИ╩ ЮМРХНЙЯХДЮМРМНЦН УЮПЮЙРЕПЮ. бЕПНЪРМН, ГЮ ЯВ╦Р ОНЯКЕДМХУ МЮ ОЕПБШУ ОНПЮУ СДЮЕРЯЪ МЕ РНКЭЙН ХГАЕФЮРЭ НЙХЯКХРЕКЭМНИ ДЕЦПЮДЮЖХХ, МН Х ЯНГДЮРЭ СЯКНБХЪ ДКЪ ЯРХЛСКЪЖХХ ОПНКХТЕПЮРХБМНЦН ОПНЖЕЯЯЮ, ЯМХГХБ БШЯНЙХИ БМЮВЮКЕ ЪБМН ╚ЖХРНРНЙЯХВЕЯЙХИ╩ БМСРПХЙКЕРНВМШИ ДХЯАЮКЮМЯ Δж (он √ юн) ДН МЕНАУНДХЛНЦН ДКЪ НЙХЯКХРЕКЭМНЦН ЛХРНЦЕМЕГЮ. нДМЮЙН Х ЩРЮ ЯРЮДХЪ Б ФХГМХ ОЕПБХВМНИ ЙСКЭРСПШ МЕ ЛНФЕР МЕ НЦПЮМХВХБЮРЭЯЪ МЕОПЕПШБМН СЦМЕРЮЧЫЕИ ЕЕ ЦХОЕПНЙЯХВЕЯЙНИ ЯПЕДНИ. б ДЮММНИ ЯХРСЮЖХХ МЮВХМЮЕР ХМЮЙРХБХПНБЮРЭЯЪ ЯЮЛ ЮДЮОРХБМШИ ЛЕУЮМХГЛ, ЯННРБЕРЯРБЕММН ЯМХФЮЕРЯЪ МЮПЮЫХБЮМХЕ ЮМРХНЙЯХДЮМРМНИ ЯХЯРЕЛШ, Ю Б ОНЯКЕДСЧЫЕЛ ОПН-ХЯУНДХР Х ПЕЦПЕЯЯХЪ ОНЯКЕДМЕИ. оПХ БШЯНЙНЛ СПНБМЕ онк, ОПЕФДЕ БЯЕЦН, ОНБПЕФДЮЧРЯЪ ЛХРНУНМДПХХ (ЯЛ. О. 1.3), ЙНКХВЕЯРБН ЙНРНПШУ ОПНДНКФЮКН АШ БНГПЮЯРЮРЭ ЙЮЙ ОПХЯОНЯНАХРЕКЭМШИ ЮЙР Б ЯКСВЮЕ ОНЯРЕОЕММНЦН ОНБШЬЕМХЪ Пн2 Б ЦЮГНБНИ ЯПЕДЕ.
мЕЯОНЯНАМНЯРЭ ЮДЮОРХБМШУ ЛЕУЮМХГЛНБ ЙКЕРЙХ Й АШЯРПНИ Х ОНКМНИ МЕИРПЮКХГЮЖХХ БМЕГЮОМН БНГМХЙЬЕИ ЦХОЕПНЙЯХХ, Я НДМНИ ЯРНПНМШ, Х БШЯНЙЮЪ СЪГБХЛНЯРЭ ЛХРНУНМДПХЮКЭМНЦН ГБЕМЮ ОПХ ОЕПНЙЯХДЮРХБМШУ ЯРПЕЯЯЮУ, Я ДПСЦНИ, НОПЕДЕКЪЧР МЕНАПЮРХЛШИ ОПНЖЕЯЯ ДЕЦЕМЕПЮЖХХ ЙКЕРНЙ ОНЯКЕ БНГМХЙМНБЕМХЪ Б МХУ ╚ЙПХРХВЕЯЙНЦН СПНБМЪ╩ ОНБПЕФДЕМХИ.
бЮФМН ГДЕЯЭ ЕЫ╦ ПЮГ НРЛЕРХРЭ: ДЕЯРПСЙРХБМШЕ ХГЛЕМЕМХЪ ХЛЕММН Б ЛХРНУНМДПХЪУ ЙЮЙ НЯМНБМШУ ОНРПЕАХРЕКЕИ н2 Х Б ЩРНЛ ЯЛШЯКЕ ЙЮЙ ЦКЮБМНИ, ЮМРХЙХЯКНПНДМНИ ЯРСОЕМХ ГЮЫХРШ Б ЮМРХНЙЯХДЮМРМНИ ЯХЯРЕЛЕ ЙКЕРЙХ МЕ НЯРЮБКЪЧР МЮДЕФД МЮ БШФХБЮМХЕ ДКЪ АНКЭЬХМЯРБЮ ЙКЕРНЙ Б ФЕЯРЙХУ СЯКНБХЪУ in vitro, ОНЯЙНКЭЙС Б ЩРНЛ ЯКСВЮЕ ПЮЯЯРПЮХБЮЕРЯЪ ЯЮЛ ЮДЮО-РХБМШИ ЛЕУЮМХГЛ ЯМХФЕМХЪ БМСРПХЙКЕРНВМНЦН Пн2 Х СПНБМЪ онк. оПХБЕДЕММШЕ ЯННАПЮФЕМХЪ ОНКМНЯРЭЧ ЯНЦКЮЯСЧРЯЪ Я ОЕПБХВМНИ ПНКЭЧ ХГЛЕМЕМХИ ЛХРНУНМ-ДПХИ Б ХМХЖХЮЖХХ ЛЕУЮМХГЛЮ ЯРЮПЕМХЪ, ОНЯРСКХПНБЮММНИ, ОПЮБДЮ, ОПХЛЕМХ-РЕКЭМН Й ЙСКЭРХБХПСЕЛШЛ in vitro ТХАПНАКЮЯРЮЛ (Kanungo, 1980).оЕПНЙЯХДЮРХБМШИ ЯРПЕЯЯ Х РНЙЯХВЕЯЙХИ ЩТТЕЙР Б СЯКНБХЪУ in vitro ЛНЦСР АШРЭ ЕЫ╦ АНКЕЕ СЯХКЕМШ, ЕЯКХ Б ЙСКЭРСПЮКЭМСЧ ЯПЕДС ББЕЯРХ ЙЮРЮКХГЮРНПШ онк, МЮОПХЛЕП ХНМШ Fe2+ ХКХ Cu2+. дЕИЯРБХРЕКЭМН, ДНАЮБКЕМХЕ Б ЯПЕДС ЙСКЭРХБХПНБЮМХЪ ЯСКЭТЮРЮ ЛЕДХ Б ЙНМЖЕМРПЮЖХХ 60 ЛЦ/К ОПХБНДХКН Й ДНЯРНБЕПМНЛС ЯМХФЕМХЧ ЯПЕДМЕИ ОПНДНКФХРЕКЭМНЯРХ ФХГМХ ЙНКНБПЮРНЙ МЮ 9 %, Ю РЮЙФЕ Й ЯСЫЕЯРБЕММН АНКЕЕ ГЮЛЕРМНЛС, ВЕЛ Б ЙНМРПНКЕ, МЮПЮЯРЮМХЧ ЙНКХВЕЯРБЮ MDA. юБРНПШ ЩРНЦН ЩЙЯОЕПХЛЕМРЮ (Enesco et al., 1989) КНЦХВМН ОНКЮЦЮЧР, ВРН ЯНЙПЮ-ЫЕМХЕ ОПНДНКФХРЕКЭМНЯРХ ФХГМХ ОПНХЯУНДХР БЯКЕДЯРБХЕ СЯЙНПЕМХЪ ХНМЮЛХ ЛЕДХ ОПНЖЕЯЯНБ ЦЕМЕПЮЖХХ ЯБНАНДМШУ ПЮДХЙЮКНБ. сЙЮГЮММЮЪ ЙНМЖЕМРПЮЖХЪ ЯСКЭТЮРЮ ЛЕДХ НЙЮГЮКЮЯЭ НОРХЛЮКЭМНИ, РЮЙ ЙЮЙ АoКЭЬХЕ (90 Х 180 ЛЦ/К) АШКХ ЯКХЬЙНЛ РНЙЯХВМШЛХ ДКЪ ЙНКНБПЮРНЙ, Ю ЛЕМЭЬЮЪ (30 ЛЦ/К) √ ЛЮКНЩТТЕЙРХБМНИ.
рЮЙХЛ НАПЮГНЛ, МЕНАПЮРХЛШЕ СЯЙНПЕММШЕ ЯРЮПЕМХЕ Х НЙХЯКХРЕКЭМЮЪ ДЕЦПЮДЮЖХЪ ЙКЕРНЙ ОПХ ПЕГЙНИ ЯЛЕМЕ ЯПЕДШ НАХРЮМХЪ Я in vivo МЮ in vitro ЪБКЪЧРЯЪ ЯКЕДЯРБХЕЛ МЕДНЯРЮРНВМНИ ЦНРНБМНЯРХ ХУ АЕГ ЯЕПЭЕГМШУ МЕЦЮРХБМШУ ОНЯКЕДЯРБХИ БНЯОПХМЪРЭ ЯРНКЭ ЙПСРНЕ СЯХКЕМХЕ ЙХЯКНПНДМНЦН БНГДЕИЯРБХЪ. еЯКХ РЮЙНИ ПЕГЙХИ ОЕПЕБНД Б МНБШЕ СЯКНБХЪ ГЮЛЕМХРЭ ╚ЛЪЦЙХЛ╩, МЮОПХЛЕП, ЛМНЦНЯРСОЕМВЮРШЛ Х ПЮЯРЪМСРШЛ БН БПЕЛЕМХ, РН ЛНФМН НФХДЮРЭ, ВРН ОПХЯСЫЮЪ ЙКЕРЙЮЛ ЯОНЯНАМНЯРЭ ЮДЮОРХПНБЮРЭЯЪ Й ОНЯРЕОЕММН МЮПЮЯРЮЧЫЕИ ЦХОЕПНЙЯХХ Б ЩРНЛ ЯКСВЮЕ ПЕЮКХГСЕРЯЪ Б ОНКМНИ ЛЕПЕ. аНКЕЕ РНЦН, Б ОПХМЖХОЕ РЮЙХЛ ЯОНЯНАНЛ ЛНФМН ДНАХРЭЯЪ ЮДЮОРЮЖХХ ЙКЕРНЙ МЕ РНКЭЙН Й НАШВМНЛС 18-21 %-МНЛС СПНБМЧ н2 Б ЮРЛНЯТЕПЕ, МН Х Й ЯСЫЕЯРБЕММН ОПЕБШЬЮЧЫХЛ ЕЦН ХЯЙСЯЯРБЕММН ЯНГДЮБЮЕЛШЛ ЦХОЕПНЙЯХВЕЯЙХЛ ЯПЕДЮЛ. б ОНДРБЕПФДЕМХЕ ЯЙЮГЮММНЛС ЯНЬКЕЛЯЪ МЮ БЕЯЭЛЮ САЕДХРЕКЭМШЕ ТЮЙРШ, ОНКСВЕММШЕ бЕКЙНЛ Я ЯНЮБР. (Valk et al., 1985). б ПЕГСКЭРЮРЕ ОНЯРЕОЕММНИ ЮДЮОРЮЖХХ Й БНГПЮЯРЮЧЫЕИ ЙНМЖЕМРПЮЖХХ н2 ХЛХ ОНКСВЕМЮ КХМХЪ ЙКЕРНЙ ЪХВМХЙЮ ЙХРЮИЯЙНЦН УНЛЪВЙЮ, СЯРНИВХБЮЪ Й БШЯНЙНЛС ЯНДЕПФЮМХЧ н2 Х ЯОНЯНАМЮЪ ОПНКХТЕПХПНБЮРЭ ДЮФЕ ОПХ 99 % н2 Б ЮРЛНЯТЕПЕ. й ЯРНКЭ ГМЮВХРЕКЭМНИ ЦХОЕПНЙЯХХ Х ГЮБХЯХЛШЛ НР МЕЕ ОПНЖЕЯЯЮЛ НЙЮГЮКХЯЭ ЮДЮОРХПНБЮММШЛХ БЯЕ ЯРСОЕМХ ГЮЫХРШ √ ЮМРХЙХЯКНПНДМЮЪ, ЮМРХПЮДХЙЮКЭМЮЪ Х ЮМРХОЕПЕЙХЯМЮЪ (ОНДПНАМЕЕ НА ЩРХУ ПЕГСКЭРЮРЮУ ЯЛ. Б ЦКЮБЕ 4).
1.8.3. хГКНФЕММШЕ ЯННАПЮФЕМХЪ НА НЯНАЕММНЯРЪУ ХГЛЕМЕМХЪ ОПННЙЯХДЮМ-РМН-ЮМРХНЙЯХДЮМРМНЦН ДХЯАЮКЮМЯЮ Б ЙСКЭРХБХПСЕЛШУ ЙКЕРЙЮУ ЙЮЙ НЯМНБМНЦН ДЕИЯРБСЧЫЕЦН ТЮЙРНПЮ ХУ ЯРЮПЕМХЪ Х РПЮМЯТНПЛЮЖХХ ЛНФМН СЯКНБМН ОПЕДЯРЮБХРЭ ЦПЮТХВЕЯЙХ (ЯЛ. ПХЯ. 11). мЮ ЙПХБНИ 1, НРПЮФЮЧЫЕИ СЙЮГЮММШЕ ХГЛЕМЕМХЪ ОПХ АШЯРПНЛ ОЕПЕЛЕЫЕМХХ ЙКЕРНЙ Б ЯПЕДС in vitro, БШДЕКЕМШ РПХ ОНЯКЕДН-БЮРЕКЭМШЕ БН БПЕЛЕМХ ЩРЮОЮ, ЙНРНПШЕ, ОНУНФЕ, ЯННРБЕРЯРБСЧР ХГБЕЯРМШЛ Б КХРЕПЮРСПЕ ЮДЮОРЮЖХНММНИ (КЮРЕМРМНИ) ТЮГЕ, ТЮГЕ КНЦЮПХТЛХВЕЯЙНЦН ПНЯРЮ Х ЯРЮЖХНМЮПМНИ ТЮГЕ. оПХ ЩРНЛ ЯРЮПЕМХЕ ЙКЕРНВМШУ ЙСКЭРСП НАШВМН ЯБЪГШБЮЕРЯЪ Я ОПНЖЕЯЯЮЛХ Б ЯРЮЖХНМЮПМНИ ТЮГЕ, ЦДЕ ЯН БПЕЛЕМЕЛ НМХ ОПЕРЕПОЕБЮЧР ПЮГКХВМШЕ ХГЛЕМЕМХЪ, ЯУНДМШЕ Я МЮАКЧДЮЧЫХЛХЯЪ Б ЙКЕРЙЮУ Б ЯНЯРЮБЕ ЯРЮПЕЧЫЕЦН НПЦЮМХГЛЮ (йЮОХРЮМНБ, 1986; уНУКНБ, 1988). б ВЮЯРМНЯРХ, ОПХ ЯРЮПЕМХХ ЙКЕРНЙ in vitro ХГЛЕМЪЧРЯЪ ТЕПЛЕМРШ, ОПНХЯУНДХР ХУ ЮМЩС- Х ОНКХОКНХДХГЮЖХЪ (Re-macle, 1989). йЮЙ Х ЙКЕРЙХ in vivo, ЙСКЭРХБХПСЕЛШЕ ЙКЕРЙХ ОН ЛЕПЕ ЯРЮПЕМХЪ МЮЙЮОКХБЮЧР КХОНТСЯЖХМНБШЕ ЦПЮМСКШ (нАСУНБЮ, щЛЮМСЩКЭ, 1984), СЙЮГШБЮЪ МЮ НВЕБХДМНЕ ОПНРЕЙЮМХЕ ОЕПЕЙХЯМШУ ОПНЖЕЯЯНБ Х НЙХЯКХРЕКЭМШЕ МЮПСЬЕМХЪ Б ЯРПСЙРСПЕ КХОХДНБ Х АЕКЙНБ. щРХ Х ПЪД ДПСЦХУ ТЮЙРНБ РЮЙ ХКХ ХМЮВЕ ЛНЦСР АШРЭ ЯНЦКЮЯНБЮМШ Я ЦХОНРЕГНИ Н ЙХЯКНПНДМН-ОЕПЕЙХЯМНЛ (ЯБНАНДМНПЮДХЙЮКЭМНЛ) ЛЕ-УЮМХГЛЕ ЯРЮПЕМХЪ. аНКЕЕ ФЕ БЯЕЦН, Б ОНКЭГС РЮЙНЦН ЛЕУЮМХГЛЮ ЯБХДЕРЕКЭЯРБСЧР ДЮММШЕ Н РНЛ, ВРН ОПХ ОНБШЬЕМХХ ЙНМЖЕМРПЮЖХХ ЮМРХНЙЯХДЮМРНБ ОПНДНКФХРЕКЭМНЯРЭ ФХГМХ ЙКЕРНЙ in vitro АНКЭЬЕ, Ю ОПХ ОНМХФЕМХХ √ ЛЕМЭЬЕ, ВЕЛ Б ЙНМРПНКЕ. рЮЙХЕ ПЕГСКЭРЮРШ ОНКСВЕМШ, МЮОПХЛЕП, ОПХ ХГЛЕМЕМХХ ЯНДЕПФЮМХЪ GSH Б ТХАПНАКЮЯРЮУ ВЕКНБЕЙЮ (Shuji, Matsuo, 1988), ЙЮРЮКЮГШ Х SOD √ Б ЙСКЭ-РХБХПСЕЛШУ МЕИПНМЮУ (Drukarch et al., 1998).
вРН ЙЮЯЮЕРЯЪ ОНКНЦХУ Х РМНЯХРЕКЭМН ОКЮБМН БНГПЮЯРЮЧЫХУ ЙПХБШУ 2 МЮ ПХЯ. 11, РН РЮЙНИ УЮПЮЙРЕП ХУ НАЗЪЯМЪЕРЯЪ РЕЛ, ВРН ГЮ ЙЮФДШЛ МЕАНКЭЬХЛ ХЯЙСЯЯРБЕММН ЯНГДЮБЮЕЛШЛ ОПХПЮЫЕМХЕЛ ОПННЙЯХДЮМРМНИ ЯНЯРЮБКЪЧЫЕИ ДХЯАЮКЮМЯЮ Δ (он √ юн) Б ЙКЕРЙЕ ЯКЕДСЕР Я МЕЙНРНПШЛ ГЮОЮГДШБЮМХЕЛ ЯННРБЕРЯРБСЧЫЕЕ ЮДЮОРХБМНЕ ОПХПЮЫЕМХЕ Б МЕИ ЮМРХНЙЯХДЮМРМНИ ЙНЛОНМЕМРШ. лМНЦНЙПЮР-МНЕ ОНБРНПЕМХЕ ЩРНИ ЮЙЖХХ Х НАЕЯОЕВХБЮЕР ОПХЯОНЯНАКЕМХЕ Х БШФХБЮЕЛНЯРЭ ЙКЕРНЙ ОПХ ОНЯРЕОЕММНЛ, ЯРСОЕМВЮРНЛ ОНБШЬЕМХХ СПНБМЪ ЦХОЕПНЙЯХХ.
б НАНХУ СЙЮГЮММШУ ЯКСВЮЪУ НАПЮРХЛ БМХЛЮМХЕ МЮ БЮПХЮМРШ, БЕДСЫХЕ Й РЮЙ МЮГШБЮЕЛНИ ╚ЯОНМРЮММНИ╩ ЛЮКХЦМХГЮЖХХ ЙКЕРНЙ (ЯЛ. ЦКЮБС 4). щРНР ТЕМНЛЕМ, Я МЮЬЕИ РНВЙХ ГПЕМХЪ, ЛНФЕР ПЕЮКХГНБШБЮРЭЯЪ КХЬЭ Б РЕУ ЙКЕРЙЮУ, ЦДЕ ДХЯАЮКЮМЯ ДНЯРХЦЮЕР ГМЮВЕМХИ Δй, СЯРНИВХБН СДНБКЕРБНПЪЧЫХУ МЕПЮБЕМЯРБС (ЯЛ. О.1.1.2)
Δо (он √ юн) < Δй (он √ юн) < Δж (он √ юн),
Ю РНВМЕЕ, Я СВЕРНЛ ╚ЮОНОРНГМШУ╩ ДХЯАЮКЮМЯНБ, √ ЯННРМНЬЕМХЧ (ЯЛ. О. 7.1.1)
Δю1 (он √ юн) < Δй (он √ юн) < Δю2 (он √ юн).
я ОНЛНЫЭЧ ОНДНАМШУ ОПНЖЕДСП, Б ЙНМЕВМНЛ ЯВЕРЕ, ТНПЛХПСЧРЯЪ ОЕПЕБХБМШЕ КХМХХ РПЮМЯТНПЛХПНБЮММШУ Х НОСУНКЕБШУ ЙКЕРНЙ, ЯОНЯНАМШУ Й ДКХРЕКЭМНЛС ЯСЫЕЯРБНБЮМХЧ БМЕ НПЦЮМХГЛЮ. б ЙНМРЕЙЯРЕ ФЕ ПЮЯЯЛЮРПХБЮЕЛШУ МЮЛХ ОПНАКЕЛ АНКЕЕ БЮФМН НОПЕДЕКХРЭЯЪ Я ОНДУНДНЛ Й ХЯЯКЕДНБЮМХЧ БГЮХЛНЯБЪГХ ЯРЮПЕМХЪ Х ЙЮМЖЕПНЦЕМЕГЮ. нДХМ ХГ МХУ, Ю ХЛЕММН ХГСВЕМХЕ ЯЮЛНЦН ОПНЖЕЯЯЮ ОНЪБКЕМХЪ НОСУНКЕБШУ ЙКЕРНЙ Б УНДЕ ЯРЮПЕМХЪ МНПЛЮКЭМШУ ЙКЕРНВМШУ ЙСКЭРСП (Witten, 1986), ОПЕДЯРЮБКЪЕРЯЪ МЮХАНКЕЕ ЕЯРЕЯРБЕММШЛ Х ОНРНЛС ОПЕДОНВРХРЕКЭМШЛ
 | ||||
| ||||
ОНДУНДНЛ. оПХ СЯРЮМНБКЕМХХ ДХЯАЮКЮМЯЮ Δ (он √ юн) Б ОПНЛЕФСРЙЕ ЛЕФДС Δй Х Δж ЙКЕРЙХ ЛНЦСР ОНДБЕПЦЮРЭЯЪ ЮОНОРНГС РХОЮ ю2 (ЯЛ. О. 7.1.1).
яНЦКЮЯМН РЕКНЛЕПМНИ РЕНПХХ, ПЕОКХЙЮРХБМНЕ ЯРЮПЕМХЕ ЙКЕРНЙ, Б РНЛ ВХЯКЕ Х Б СЯКНБХЪУ in vitro, ЯБЪГЮМН Я СЙНПНВЕМХЕЛ РЕКНЛЕП ОНЯКЕ ЙЮФДНЦН ЛХРНГЮ, БОКНРЭ ДН МЕЙНРНПНИ ЛХМХЛЮКЭМНИ ДКХМШ, ПЕГСКЭРЮРНЛ ВЕЦН ЪБКЪЕРЯЪ СРПЮРЮ РЮЙХЛХ ЙКЕРЙЮЛХ ЯОНЯНАМНЯРХ Й ДЕКЕМХЧ (ЯЛ. О. 1.4.3 Х 1.4.4). юМЮКХГ ХГБЕЯРМНИ КХРЕПЮРСПШ ОН ЩРНЛС БНОПНЯС ОНЙЮГШБЮЕР, ВРН ДЮММШИ ОНЯРСКЮР Б МЕЙНРНПШУ ЯКСВЮЪУ МЕ ОНДРБЕПФДЮЕРЯЪ. оПХЛЕПНЛ РНЛС ЯКСФЮР ХЯЯКЕДНБЮМХЪ йЮПЛЮМЮ Х ЯНЮБР. (Carman et al., 1998), ОПНБЕДЕММШЕ МЮ ДХОКНХДМШУ ЩЛАПХНМЮКЭМШУ ЙКЕ-РЙЮУ ЯХПХИЯЙНЦН УНЛЪВЙЮ (SHE). щРХ ЙКЕРЙХ ОНЯКЕ 20-30 ЖХЙКНБ СДБНЕМХЪ ОПЕЙПЮЫЮКХ ОПНКХТЕПЮЖХЧ, РЕПЪКХ ЯОНЯНАМНЯРЭ БЯРСОЮРЭ Б S-ТЮГС ОНЯКЕ ЯРХЛСКЪЖХХ ЯШБНПНРЙНИ. б РН ФЕ БПЕЛЪ ЙКЕРЙХ SHE ЩЙЯОПЕЯЯХПНБЮКХ РЕКНЛЕПЮГС Б РЕВЕМХЕ БЯЕЦН ПЕОКХЙЮРХБМНЦН ФХГМЕММНЦН ЖХЙКЮ, Ю ЯПЕДМХИ ПЮГЛЕП РЕКНЛЕП МЕ СЛЕМЭЬЮКЯЪ. бШУНДХР, ВРН in vitro ЙКЕРЙХ ЛНЦСР ХМНЦДЮ ЯРЮПЕРЭ ОН ЛЕУЮМХГЛЮЛ, МЕ ЯБЪГЮММШЛ Я ОНРЕПЕИ РЕКНЛЕП.
йЮЙ ОПЕДЯРЮБКЪЕРЯЪ МЮЛ, Б СЙЮГЮММНЛ ЯКСВЮЕ ЯБНХ ЙНППЕЙРХБШ БМНЯЪР СЯКНБХЪ ЦХОЕПНЙЯХХ Б ЯПЕДЕ ЙСКЭРХБХПНБЮМХЪ. еЯКХ Б ЯНЯРНЪМХХ СЛЕПЕММН ОНБШЬЕММНЦН СПНБМЪ ютй Х ОЕПНЙЯХДЮЖХЪ БШОНКМЪЧР МЕПЕДЙН ОНГХРХБМШЕ ТСМЙЖХХ, ЮЙРХБХПСЪ НРДЕКЭМШЕ ЩРЮОШ ОПНУНФДЕМХЪ ЛХРНЦЕММНЦН ЯХЦМЮКЮ, ПЕОКХЙЮ-ЖХХ, РПЮМЯЙПХОЖХХ Х ХМШУ ОПНЖЕЯЯНБ (НА ЩРНЛ ЦНБНПХКНЯЭ Б ПЪДЕ ОПЕДШДСЫХУ ОЮПЮЦПЮТНБ Х СОНЛХМЮЕРЯЪ Б МЕЙНРНПШУ ОНЯКЕДСЧЫХУ), РН Б ЯКСВЮЕ ХМРЕМЯХБМНЦН НЙХЯКХРЕКЭМНЦН ЯРПЕЯЯЮ МЕХГАЕФМШ Х МЕЦЮРХБМШЕ ОНЯКЕДЯРБХЪ. мЮОПХЛЕП, ВЮЯРЭ ЛЮЙПНЛНКЕЙСК, Б РНЛ ВХЯКЕ СВЮЯРБСЧЫХУ Б ЛХРНЦЕМЕГЕ, ЛНФЕР АШРЭ ЛНДХТХЖХПНБЮМЮ, ВРН, МЕГЮБХЯХЛН НР ЮЙРХБМНЯРХ РЕКНЛЕПЮГШ Х ДКХМШ РЕКНЛЕП, ДНК-ФМН ХМЦХАХПНБЮРЭ ОПНКХТЕПЮЖХЧ Х/ХКХ ХМДСЖХПНБЮРЭ ЙЮЙХЕ-РН ДПСЦХЕ МЮПСЬЕМХЪ, БОКНРЭ ДН ОПХБНДЪЫХУ Й ЦХАЕКХ ЙКЕРНЙ.
йЮЙ АШ РЮЛ МХ АШКН, ДБЕ ОПХВХМШ ЯРЮПЕМХЪ ЙКЕРНЙ in vitro √ МЮЙНОКЕМХЕ НЬХАНЙ Б СЯКНБХЪУ ЯНДЕПФЮМХЪ ХУ Б ЙСКЭРСПЕ Х СЙНПНВЕМХЕ РЕКНЛЕП √ НЯРЮЧРЯЪ БЯЕ ФЕ МЮХАНКЕЕ БЕПНЪРМШЛХ. оНКЮЦЮЧР, ВРН Б НАНХУ ЯКСВЮЪУ ЮЙРХБХПСЧРЯЪ ЯХЯРЕЛШ АЕКЙНБ П53 Х Rb, Ю ОПХ МЮПСЬЕМХХ ХУ ТСМЙЖХХ ОПНХЯУНДХР РПЮМЯТНПЛЮЖХЪ ЙКЕРНЙ (Sherr, DePinho, 2000). б АНКЕЕ НАЫЕЛ ОКЮМЕ МЮЛ БХДХРЯЪ ЯКЕДСЧЫЕЕ: Б РНЙЯХВЕЯЙХУ ЦХОЕПНЙЯХВЕЯЙХУ СЯКНБХЪУ ЙСКЭРХБХПНБЮМХЪ МНПЛЮ-КЭМШЕ ЙКЕРЙХ, ЯРЮПЕЪ, ОПЕРЕПОЕБЮЧР, ЯЙНПЕЕ БЯЕЦН, ЮОНОРНГ ю1, Ю НОСУНКЕБШЕ ЙКЕРЙХ √ ЮОНОРНГ ю2. б ЯКСВЮЕ МЕОНКЮДНЙ Б ЛЕУЮМХГЛЕ ЮОНОРНГЮ ОЕПБШЕ ОНДБЕПЦЮЧРЯЪ МЕНОКЮЯРХВЕЯЙНИ РПЮМЯТНПЛЮЖХХ, БРНПШЕ ФЕ √ НЙХЯКХРЕКЭМНЛС ЖХРНКХГС (ЯЛ. О. 7.1.1).
дНОНКМХРЕКЭМНИ ОПХВХМНИ, ЯОНЯНАЯРБСЧЫЕИ СЯХКЕМХЧ ОПНЖЕЯЯНБ НЙХЯКХРЕКЭМНИ ДЕЯРПСЙЖХХ Б ЙКЕРЙЮУ in vitro, ЛНФЕР ЯВХРЮРЭЯЪ Х РЕОКН, ЙЮЙ ОНЯРНЪММН ДЕИЯРБСЧЫХИ ТЮЙРНП НЙПСФЮЧЫЕИ ЯПЕДШ. дЕИЯРБХРЕКЭМН, Я ОНЛНЫЭЧ БШЯНЙН-ВСБЯРБХРЕКЭМНЦН ЛЕРНДЮ (НОХЯЮМХЕ ЕЦН ОПХБНДЪР ЮБРНПШ аПСЯЙНБ Х ДП., 2001) АШКН ОНЙЮГЮМН, ВРН ОНД ДЕИЯРБХЕЛ РЕОКЮ Б БНДМШУ ПЮЯРБНПЮУ ЦЕМЕПХПСЧРЯЪ ютй. б ПЕГСКЭРЮРЕ РЕОКНБНИ ЮЙРХБЮЖХХ ПЮЯРБНПЕММНЦН Б БНДЕ ЮРЛНЯТЕПМНЦН н2 ОПНРЕЙЮЕР ОНЯКЕДНБЮРЕКЭМНЯРЭ ПЕЮЙЖХИ
н2 → 1н2 → н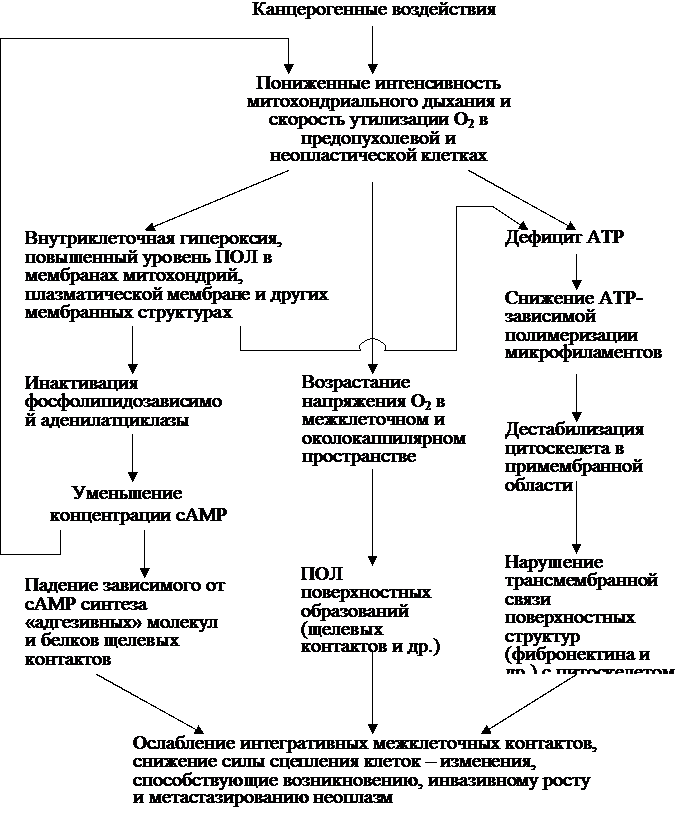 → мн2˙ → м2н2 → нм˙.
→ мн2˙ → м2н2 → нм˙.
нАПЮГСЧЫХЕЯЪ ютй, ОН-БХДХЛНЛС, Х ЯНДЕИЯРБСЧР РЕОКНБНЛС ОНБПЕФДЕМХЧ дмй Х ДПСЦХУ АХНКНЦХВЕЯЙХУ ЛНКЕЙСК ОСР╦Л ХУ ╚ЮБРННЙХЯКЕМХЪ╩.
мЮЙНМЕЖ, НРЛЕРХЛ ЕЫЕ НДХМ ЯОНЯНА ХМРЕМЯХТХЙЮЖХХ ОПНЖЕЯЯЮ ЯРЮПЕМХЪ ЙКЕРНЙ Б СЯКНБХЪУ in vitro Я ОНЛНЫЭЧ ОПНЖЕДСПШ ЮМНЙЯХХ √ ПЕНЙЯХЦЕМЮЖХХ, ПЕГСКЭРЮРШ ЙНРНПНИ, ОН МЮЬЕЛС ЛМЕМХЧ, МЮХАНКЕЕ БШПЮФЕМН НРПЮФЮЧР ЯСРЭ ЙХЯКНПНДМН-ОЕПЕЙХЯМНИ ЛНДЕКХ ЯРЮПЕМХЪ. нЯМНБС ЛЕУЮМХГЛЮ ЯРЮПЕМХЪ Б ДЮММНЛ ЯКСВЮЕ ЯНЯРЮБКЪЧР ДБЮ ОПХМЖХОХЮКЭМШУ ЩТТЕЙРЮ: ЮДЮОРХБМНЕ ЯНЙПЮЫЕМХЕ (НЯКЮАКЕМХЕ) ЛХРНУНМДПХЮКЭМНИ АЮГШ Б ОЕПХНД ЮМНЙЯХХ ХКХ ЦХОНЙЯХХ (ЯЛ. БШЬЕ); ГМЮВХРЕКЭМНЕ СЯХКЕМХЕ онк Х ДПСЦХУ ОПНЖЕЯЯНБ НЙХЯКХРЕКЭМНИ ДЕЯРПСЙЖХХ ОПХ ОНЯКЕДСЧЫЕИ ПЕНЙЯХЦЕМЮЖХХ БЯКЕДЯРБХЕ ПЕГЙНЦН ОНБШЬЕМХЪ Пн2 (НРМНЯХРЕКЭМН ЯНЯРНЪМХЪ ЮМНЙЯХХ) Х МЕБНГЛНФМНЯРХ АШЯРПНИ СРХКХГЮЖХХ ХГАШРНВМНЦН н2 ╚ЮМНЙЯХВЕЯЙХЛХ╩ ЛХРНУНМДПХЪЛХ. яРЕОЕМЭ ОЕПНЙЯХДЮРХБМНЦН ЯРПЕЯЯЮ Х, ЯКЕ-ДНБЮРЕКЭМН, ЯЙНПНЯРЭ ЯРЮПЕМХЪ ЙКЕРНЙ АСДСР ГЮБХЯЕРЭ НР ДКХРЕКЭМНЯРХ ОПЕАШБЮМХЪ ХУ Б ЯНЯРНЪМХХ ЮМНЙЯХХ: ВЕЛ ОПНДНКФХРЕКЭМЕЕ ЩРНР ОЕПХНД, РЕЛ КСВЬЕ ЯЛНФЕР ЮДЮОРХПНБЮРЭЯЪ ЛХРНУНМДПХЮКЭМЮЪ АЮГЮ Й МХГЙНЛС СПНБМЧ Пн2 Х РЕЛ ГМЮВХРЕКЭМЕЕ АСДЕР СПНМ ЙКЕРНЙ ОНЯКЕ СЯРПЮМЕМХЪ ХЬЕЛХХ.
оПХЛЕПНЛ ПЕЮКХГЮЖХХ ЯРЮПЕМХЪ ЙКЕРНЙ ОН СЙЮГЮММНЛС ╚ЯЖЕМЮПХЧ╩ ЛНФЕР ЯКСФХРЭ ЯКЕДСЧЫХИ ТЮЙР. цЕОЮРНЖХРШ, БШДЕКЕММШЕ С ЙПШЯ ПЮГМНЦН БНГПЮЯРЮ, ОНДБЕПЦЮКХ 2-ВЮЯНБНИ ЮМНЙЯХХ Х 1-ВЮЯНБНИ ПЕНЙЯХЦЕМЮЖХХ. сЯРЮМНБКЕМН, ВРН Б ПЕНЙЯХЦЕМЮЖХНММНИ ТЮГЕ ЦЕОЮРНЖХРШ ОПНДСЖХПСЧР АНКЭЬНЕ ЙНКХВЕЯРБН ЙХЯКНПНДМШУ ПЮДХЙЮКНБ, НРБЕРЯРБЕММШУ ГЮ ОНБПЕФДЕМХЕ ХУ ЛЕЛАПЮМ Х ГЮ ДПСЦХЕ ОПХ-ВЮЯРМШЕ Й ЯРЮПЕМХЧ ЯРПСЙРСПМН-ТСМЙЖХНМЮКЭМШЕ ХГЛЕМЕМХЪ, ОПХВЕЛ ЯРЮПШЕ ЙКЕРЙХ АШКХ ВСБЯРБХРЕКЭМЕЕ Й ПЕОЕПТСГХНММНИ РПЮБЛЕ (Gasbarrini et al., 1998). оНДНАМНЦН ПНДЮ ТЮЙРШ ПЮЯЯЛЮРПХБЮЧРЯЪ МЮЛХ Х Б ЦКЮБЕ 4 Б ЯБЪГХ Я НАЯСФДЕ-МХЕЛ ЛЕУЮМХГЛЮ ЯРЮПЕМХЪ Х ╚ЯОНМРЮММНИ╩ ЛЮКХЦМХГЮЖХХ ЙКЕРНЙ Б ЙСКЭРСПЕ.
еЫЕ ОН РЕЛЕ 1.8. яРЮПЕМХЕ ЙКЕРНЙ in vitro:
- яБНИЯРБЮ НОСУНКЕБШУ ЙКЕРНЙ in vitro
- ЯСЫЕЯРБНБЮМХЕ ОСРЕИ СЙНПНВЕМХЪ РЕКНЛЕП ОПХ ЯРЮПЕМХХ ЙКЕРНЙ
- хГЛЕМЕМХЪ ЙКЕРНЙ ОПХ ЯРЮПЕМХХ.
- 1.2.2. нДМХЛ ХГ МЮХАНКЕЕ НВЕБХДМШУ Х ГЮЛЕРМШУ ХГЛЕМЕМХИ ЙКЕРНЙ ОПХ ЯРЮПЕМХХ ФХБШУ НПЦЮМХГЛНБ
- хГСВЕМХЕ in vitro ХГЛЕМЕМХЪ ЯХМРЕРХВЕЯЙНИ ЮЙРХБМНЯРХ НОСУНКЕБШУ ЙКЕРНЙ ЮЯЖХРМНЦН БЮПХЮМРЮ ПЮЙЮ ЪХВМХЙНБ ЙПШЯ ОПХ ДЕИЯРБХХ оЕло Х БХМЙПХЯРХМЮ
- хГСВЕМХЕ in vitro ЯХМРЕРХВЕЯЙНИ ЮЙРХБМНЯРХ ЙКЕРНЙ КХЛТШ АНКЭМШУ ПЮЙНЛ ЪХВМХЙНБ ОПХ ДЕИЯРБХХ оЕло, Ю РЮЙФЕ ДНЙЯНПСАХЖХМЮ Б ЯНВЕРЮМХХ Я ЩОХРЮКЮЛХМНЛ Х ПХУКНЙЮХМНЛ
- яРЮПЕМХЕ Х АНКЕГМХ
- хГЛЕМЕМХЪ Б НПЦЮМХГЛЕ ОПХ ЯРЮПЕМХХ
- 1.5. яРЮПЕМХЕ ЯХЯРЕЛ НПЦЮМХГЛЮ
- оСРХ БНГДЕИЯРБХЪ МЮ ЯРЮПЕМХЕ
- хГЛЕМЕМХЪ ЯХЯРЕЛ ОПХ ЯРЮПЕМХХ.
- 1.6. мЕЙНРНПШЕ ╚ОЮПЮДНЙЯЮКЭМШЕ╩ ТЕМНЛЕМШ ОПХ ЯРЮПЕМХХ
- 1.4.5. бМСРПХЙКЕРНВМЮЪ ЦХОЕПНЙЯХЪ ЙЮЙ ПЕГСКЭРЮР ОЕПБХВМНЦН ОПНЖЕЯЯЮ ЯРЮПЕМХЪ ЛХРНУНМДПХИ
- НДХМ ХГ НЯМНБМШУ ЙНЛОНМЕМРНБ, НРБЕРЯРБЕММШУ ГЮ ЯРЮПЕМХЕ,
- оПХВХМШ Х ЛЕУЮМХГЛШ ЯРЮПЕМХЪ
- 1.6 вСБЯРБХРЕКЭМНЯРЭ ЛХЙПНЛХЖЕРНБ √ БНГАСДХРЕКЕИ НМХУНЛХЙНГЮ in vitro
- яРЮПЕМХЕ
- 1.5.3. мЮПСЬЕМХЪ Б ЯХЯРЕЛЕ ХЛЛСМХРЕРЮ ОПХ ЯРЮПЕМХХ
- цЕРЕПНЦЕММНЯРЭ ЯРЮПЕМХЪ
- цКЮБЮ 9. яРЮПЕМХЕ