2.1. Митохондрии как первичный объект повреждения ииндукции пероксидативного состояния в клетке
Как уже отмечалось в п.1.1.1, в ответ на рост рО2 в земной атмосфере живая природа реализовала систему защитных механизмов, предохраняющих клетку от избыточного накопления в ней токсических продуктов ПОЛ.
Антиоксидант-ная система защиты, по общему мнению исследователей (Барабой и др., 1992; Gutteridge, 1995 и др.), является иерархической и осуществляется не менее чем на трёх уровнях (Лю, Ефимов, 1976; Лю, Шайхутдинов, 1991).Первая и основная ступень защиты – антикислородная. За счёт активности дыхательных ферментов и специальной группы соединений, депонирующих избыточный О2, данная ступень поддерживает внутри клеток довольно низкие значения рО2, порядка 1-5 мм рт. ст., достаточные, однако, для тканевого дыхания и энергообеспечения. Выводы о глубокой внутриклеточной гипоксии в большинстве нормальных тканей с достаточно высокой оксигенацией внеклеточного пространства считаются надёжными, так как однозначные результаты получены различными методами и в независимых исследованиях (Ярмоненко, Эпштейн, 1977). Действительно, в ряде последующих работ подтверждено, что измеренное в цитозоле клеток (кардиомиоциты, гепатоциты и др.) значение рО2 составляет всего 0,4-4,0 мм рт. ст. (Gaeski, Honig, 1986; Makiguchi et al., 1987). Непосредственно же около митохондрий предположительно рО2 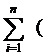 1 мм рт. ст., а в их матриксе – 0,01-0,1 мм рт. ст., т.е. на уровне критического рО2 для цитохромоксидазы как терминального фермента дыхательной цепи (Иванов, 1996). По ещё одним сведениям (см. Скулачев, 2001), цитохромоксидаза может восстанавливать О2 даже при в 100 раз меньших его концентрациях, чем в воде при нормальном атмосферном давлении.
1 мм рт. ст., а в их матриксе – 0,01-0,1 мм рт. ст., т.е. на уровне критического рО2 для цитохромоксидазы как терминального фермента дыхательной цепи (Иванов, 1996). По ещё одним сведениям (см. Скулачев, 2001), цитохромоксидаза может восстанавливать О2 даже при в 100 раз меньших его концентрациях, чем в воде при нормальном атмосферном давлении.
Анткислородная линия защиты не в состоянии, вероятно, полностью предотвратить возможные негативные последствия избыточного ПОЛ, поскольку необходимые для него свободные радикалы образуются в процессах нормального метаболизма и индуцируются различными агентами.
Кстати, особенности электронной структуры молекулы О2, выступающей в роли акцептора электронов, предопределяют синтез О , Н2О2 и НО˙ в качестве нормальных промежуточных продуктов превращения О2 + 4 е + 4 Н+ → 2 Н2О. Эта реакция складывается фактически из 4-х одноэлектронных переходов, в ходе которых и порождаются указанные АФК (Бохински, 1987):
, Н2О2 и НО˙ в качестве нормальных промежуточных продуктов превращения О2 + 4 е + 4 Н+ → 2 Н2О. Эта реакция складывается фактически из 4-х одноэлектронных переходов, в ходе которых и порождаются указанные АФК (Бохински, 1987):О2 + 1 е → О;
О + 1 е + 2 Н+ → Н2О2;
Н2О2 + 1 е + Н+ → Н3О2 → Н2О + НО˙;
НО˙ + 1 е + Н+ → Н2О

 : О2 + 4 е + 4 Н+ → 2 Н 2О.
: О2 + 4 е + 4 Н+ → 2 Н 2О.
Поэтому существуют последующие более «тонкие» ступени защиты – анти-радикальная и антиперекисная, надёжная работа которых зависит от исправного функционирования антикислородной линии защиты. При безупречном действии последней роль третьей из названных ступеней может даже казаться незначительной или действительно быть таковой. Примером тому служат мыши с недостаточностью клеточной GPX, которые развиваются нормально, не обнаруживая повышенную чувствительность к гипероксии. У таких мышей не выявлено каких-либо изменений в утилизации экзогенной Н2О2, содержании карбонильных групп в белках и пероксидации липидов (Ho et al., 1997).
С другой стороны, дефекты в антикислородной линии защиты могут определять гипероксию и пероксидацию в клетке, неэффективность всей анти-оксидантной системы. Правда, на случаи возникновения подобных ситуаций природой, похоже, предусмотрен механизм усиления последующей антипере-кисной ступени, как бы нацеливающей последнюю на возрастающую на неё нагрузку. Принципиально такая возможность, на наш взгляд, заложена сущес-твованием в 5´-фланкирующей области гена GPX человека так называемого элемента реакции на О2 – ORE1, а также нескольких копий сайтов связывания фактора транскрипции АР-1 (Jornot, Jurnod, 1997).
Здесь важно выяснить, на какой диапазон изменений рО2 «рассчитана» реакция указанного элемента. Авторы цитируемой работы, исследуя на культуре эндотелиальных клеток механизмы ORE1-опосредованной индукции гена GPX в условиях гипероксии, установили: при повышении содержания О2 (до 95 %) в атмосфере увеличи-ваются скорость транскрипции гена GPX и концентрация мРНК-GPX. Однако само действие и (или) эффективность такого механизма при реально возможных in vivo значениях внутриклеточной гипероксии требуют ещё доказательств.2.1.1. Среди внутриклеточных органелл и многих биополимеров, подвергающихся воздействию канцерогенных факторов, изменение лишь некоторых из них ответственно за опухолевые последствия. Реальным первичным объектом такого воздействия с указанным исходом представляются, прежде всего, митохондрии – основные потребители О2 в клетке. Обоснованию этой точки зрения в своё время был посвящён ряд обстоятельных обзорно-теоретических работ (Wenner, Tomei, 1981; Игнатова, 1984). «Митохондриальной» концепции на основе собственных систематических данных придерживается и Сигер (Seeger, 1985). Им показано, что в опухолевых клетках повреждены мембраны митохондрий и разрушена дыхательная цепь, окислительные процессы снижены и преобладают процессы брожения; любые липофильные вещества, способные вызывать повреждение митохондрий и инактивацию цитохромов, приводят к злокачественной трансформации клеточных культур. По данной логике, экзогенные ингибиторы дыхания также представляют собой канцерогены, непосредственно снижающие эффективность антикислородной ступени защиты. Действительно, в отношении ротенона – известного ингибитора митохондриального транспорта электронов – имеются прямые доказательства индукции им опухолей (Gosalvez, 1983). Но если объектом приложения того же роте-нона и других подобных ему ингибиторов, а также экзогенных прооксидантов окажутся уже состоявшиеся опухолевые клетки, то последние, скорее всего, подвергнутся апоптозу типа А2 или окислительному цитолизу (см.
п.7.1) вследствие установления в них более высоких, чем до воздействия указан-ных агентов, уровней АФК и ПОЛ. Отдельные факты такого рода известны (Теплова и др., 1999).Несколько раньше убедительные литературные и собственные факты о воздействии на митохондрии химических канцерогенов различных классов представили Кобляков и Рябых (1977). Уже на ранних стадиях гепатоканце-рогенеза ими обнаружены такие эффекты, как торможение дыхания и нару-шение структуры митохондрий. В этом отношении особенно показательны следующие данные (Boitier et al., 1995). Через 19 мес после применения диэтилнитрозамина в митохондриях гепатомы складываются выраженный дефицит комплекса I и снижение активности комплекса III дыхательной цепи. Одновременно в этих органеллах снижается активность антиоксидантных ферментов, а концентрации О и Н2О2 соответственно повышаются. Ответственными за инактивацию комплексов I и III дыхательной цепи являются, предположи-тельно, супероксидные радикалы.
Чен с соавт. (Chen et al., 1984), сравнивая различные параметры митохондрий в опухолевых и нормальных клетках культур различных линий, выявили значительные различия по одному или нескольким признакам: числу, длине, размеру, морфологии, распределению, подвижности, мембранному потенциалу, градиенту рН и др. По некоторым из них оценивались жизненные функции митохондрий и нарушения механизма дыхания у опухолевых клеток, а такие изменения митохондрий, как снижение их числа, дефектное дыхание, изменение топографии рассматриваются ими в качестве сопутствующих опу-холевой трансформации. На уменьшение количества и размера митохондрий, упрощение их ультраструктуры и полиморфность в опухолевой клетке по сравнению с нормальной ранее указывали и другие исследователи (Шапошников, Киселев, 1978; Голубев, 1981; Azgiles, Azcon-Bieto, 1988), причём снижение мощности митохондриальной системы представляют как коканцерогенный фон, способствующий малигнизации клеток и, возможно, приводящий к ней (Гобеев и др., 1978).
Согласно материалам международного симпозиума (Амстердам, 1985), связь между митохондриями, ядром и клеточной мембраной вновь поднимает старый вопрос об участии митохондрий в канцерогенезе.
Более того, концепция использования при химиотерапии противомитохондриальных веществ основана на представлении о том, что митохондрии в раковых клетках дефектны. В последующие годы корреляция канцерогенеза с недостаточностью митохон-дриального дыхания подтверждалась неоднократно. Так, в исследованных гли-омах человека выявлены изменения структуры крист и плотности матрикса митохондрий, а переключение энергетического метаболизма в глиомах на гликолиз связывается с низким содержанием в них нормально функционирующих митохондрий (Oudard et al., 1997). В ряде других работ обобщены данные о снижении в опухолевых клетках количества и качества митохондрий, рассмотрены различные аспекты канцерогенеза, включая мутагенез, поддержание озлокачествлённого фенотипа и участие в этих процессах митохондрий (см., например, Capuano et al., 1997; Cavalli, Liang, 1998; Reed, 1999). К разряду подобных событий относится и то, что мутации в гене SDHD, кодирующем один из белков комплекса II дыхательной цепи митохондрий, играют важную роль в патогенезе определённых опухолей, в частности, при наследственной паранглиоме, которая характеризуется развитием доброкачественных васкуляризованных опухолей головы и шеи (Baysal et al., 2000).Внутри митохондрий объектом прямой атаки некоторых канцерогенных факторов могут быть не только дыхательные ферменты, но и ДНК Последняя, как известно, кодирует ряд необходимых для сборки и функционирования митохондрий белков, в том числе ферментов дыхательной цепи. В результате подавляются синтез ряда митохондриальных цитохромов, транспорт электронов и окислительное фосфорилирование. Так, полициклические ароматические соединения разных канцерогенных потенций связываются с мтДНК в десятки и сотни раз больше, чем с ядерной (Wilkie et al., 1983). Индекс ковалентного свя-зывания метаболитов канцерогенного гексахлорбутадиена для яДНК из почек мышей, получавших данный препарат, составляет 27, а для ДНК из печени – всего 6. Для мтДНК же из почек и печени тех же животных этот индекс очень высок и равен соответственно 7500 и 500 (Schrenk, Dekant, 1989).
Одна из причин подобной предпочтительности состоит в том, что «вилки репликации мтДНК как наиболее уязвимое место для воздействия канцерогенов и митогенов присутствуют в клетках на протяжении всего клеточного цикла, что не наблюдается для репликативных вилок в ядре» (Игнатова, 1984). Кроме того, яДНК менее уязвима для повреждающих факторов вследствие защищённости различными белками и суперскрученности.Считают также, что мтДНК более чувствительна к химическим канцеро-генам из-за окруженности её липидсодержащей мембраной, способной акку-мулировать липофильные канцерогены, в частности 3,4-бензпирен. Более того, митохондриальные ДНК, ДНК- и РНК-полимеразы тесно ассоциированы с этой мембраной и потому могут подвергаться воздействию накапливаемых в ней канцерогенов, что приведёт к нарушению синтеза и транскрипции мтДНК (Виленчик, 1985). Подробнее эффекты взаимодействия химических канцерогенов с митохондриями рассмотрены в п.6.2 нашей предыдущей монографии (Лю, Шайхутдинов, 1991).
Известны факты высокой чувствительности мтДНК и к некоторым лечеб-ным химиопрепаратам, Например, после воздействия на клетки яичников китайского хомячка цисплатином содержание аддуктов в мтДНК почти на порядок превышало таковое в яДНК. Кроме того, в мтДНК в отличие от яДНК отсутствовала репарация. Количественные оценки аддуктов получены с помощью иммунологического детектирования ДНК и подтверждены методом электронной микроскопии с коллоидным золотом (Olivero et al., 1996). Чувстви-тельность мтДНК к канцерогенам и антиканцерогенам, многократно превышающая таковую яДНК, установлена и в ряде других работ (Ames et al., 1993; King, 1995).
По указанным выше причинам экспрессия митохондриальных генов, в том числе кодирующих компоненты системы окислительного фосфорилирования, в опухолях нарушается. Например, по данным Нозерн-блотинга, в опухолях почек различных типов и в полученных из них стабильных клеточных линиях стационарная концентрация мРНК – транскриптов мтДНК снижена по срав-нению с нормальной почечной тканью (Faure-Vigny et al., 1996). С другой стороны, отмечаются факты модификации ядерного генома опухолей фраг-ментами мтДНК. Так, исследование блот-гибридизацией по Саузерну мтДНК-подобных вставок в яДНК показало, что содержание и мобильность их в опухолях мыши и крысы существенно выше, чем в гомологичных нормальных тканях (Hadler et al., 1998).
Некоторые из ферментов митохондрий в клетках неоплазмы (цитохром с, цитохромоксидаза, сукцинатдегидрогеназа, малатдегидрогеназа и др.) характе-ризуются, как правило, пониженной активностью (Дмитриева, 1977; Гобеев, Березов, 1979 и др.). Причины падения активности этих ферментов разные, но главные – это их прямая или косвенная инактивация, а также уменьшение содержания некоторых из них в митохондриях. Особого внимания среди дыха-тельных энзимов заслуживает терминальная цитохромоксидаза. В составе электроно-транспортной цепи она «работает» на антикислородной линии защиты, способствуя установлению низких значений внутриклеточного рО2. А по некоторым данным (Налбадян, 1984), цитохромоксидаза обладает и SOD-актив-ностью. Перехватывая О, она защищает в своём микроокружении «собственные» липиды в реакции ПОЛ. Торможение процессов ПОЛ возрастает при увеличении концентрации цитохромоксидазы и полностью устраняется различными её ингибиторами. По этим фактам цитохромоксидаза выполняет, по существу, двойную антиоксидантную функцию на двух первых ступенях защиты. Поэтому инактивация или повреждение её в составе антиоксидантной системы должны быть особенно «чувствительными» с точки зрения возможных пероксидативных последствий в клетке.
Нарушение антикислородной ступени защиты может начаться уже на стадии «митохондриального» старения клетки (см. п.1.3) и затем в ускоренном темпе продолжиться при трансформации её в опухолевую под влиянием канцерогенных факторов. К этому нарушению, возможно, причастны белковые ингибиторы дыхания, которые, по данным ряда исследований, синтезируются опухолевой клеткой. Они влияют на энергетические процессы нормальных клеток, угнетая в них дыхание и потребление О2, т.е. создают энергетическую ситуацию, наблюдаемую в опухолях. Продолжая исследование природы обнаруженных ингибиторов дыхательных белков, Рапопорт с соавт. (Rapoport et al., 1979) установили их идентичность липооксигеназе (LOX), выделенной из ретикулоцитов крови анемичных кроликов. В присутствии этого фермента одновременно с LOX-действием происходит необратимая инактивация некоторых SH-ферментов. По данным этих авторов, существуют LOXs другого типа в лёгких, селезёнке, почках и эпителиальных опухолях. Механизм подавления LOX дыхательных ферментов тогда окончательно не был установлен.
Позже той же исследовательской группой показано, что LOX обратимо ингибирует цитохром-с-оксидазу в митохондриях и субмитохондриальных частицах сердца быка путём химической модификации фосфолипидов (Rapoport, Schewe, 1986), активация же митохондриальной 12-LOX ведёт к дезынтеграции митохондрий при созревании ретикулоцитов в эритроциты (Schnurr et al., 1996). Более того, было сформулировано общее положение об участии LOX в программированной деградации разных внутриклеточных органелл в процессе дифференцировки не только ретикулоцитов, но и ряда других специализи-рованных типов клеток (Van Leyen et al., 1998). Перед началом деградации органелл LOX интегрируется в их мембраны и, увеличивая проницаемость, способствует высвобождению содержимого органелл, доступности их белков для действия протеаз.
Приведённые выше убедительные факты снижения количества и качества митохондрий в опухолевых клетках объясняют сравнительно низкую энергети-ческую эффективность их дыхания по сравнению с таковой клеток гомологич-ных нормальных тканей (Голубев, 1981; Chen et al., 1984; Seeger, 1985; Azgiles, Azcon-Bieto, 1988; Schmidt et al., 1991; Baggeto, 1992; Luciakova, Kuzela, 1992; Capuano et al., 1997 и др.). Отрицать это принципиально важное изменение и его последствия для судеб соответствующих клеток, считая их ошибочными, по крайней мере, не логично. К тому же, следует иметь в виду, что данные сдвиги объективно начинают возникать уже в доканцерогенезный период, на стадии старения клеток (см. главу 1), более выражено проявляясь в ходе и после их трансформации.
Таким образом, доказательность многих исследований о недостаточности окислительного фосфорилирования в митохондриях опухолевых клеток не вы-зывает сомнений. Тем не менее, иногда гипотезы с противоположным мнением продолжают высказываться. Например, Рэй и Рэй (Ray, Ray, 1997), признавая резкое изменение в злокачественных клетках части дыхательной цепи (митохондриального комплекса I), полагают всё же, что именно это обстоятельство приводит к избыточному образованию АТР в указанных клетках и создаёт тем самым необходимые энергетические условия для их роста, размножения и окончательного становления свойств злокачественности. Следует, однако, приз-нать, что повышение продукции АФК при канцерогенезе через первичное повреждение митохондриального дыхания есть, хотя и главный, но не единст-венный путь. Некоторые канцерогены сами являются источником ферментатив-ного возникновения из них АФК (О). К таковым относится, в частности, 4-нитрохинолин-N-оксид, причём образование аддукта супероксидного радикала, 5,5-диметил-1-пирролин-N-оксида, ингибируется SOD (Fann et al., 1999).
Приведённым выше фактам, вообще говоря, противоречат данные о том, что в клетках различных опухолей (лимфомы, гепатомы, рака предстательной железы и др.) наряду со снижением числа митохондрий и активности дыхательных ферментов возрастает экспрессия кодирующих их митохондриальных генов. Подробная информация на этот счёт приведена недавно в статье Николаева с соавт. (2001) и в цитируемых ими работах. В частности, отмечается повышение уровня экспрессии генов субъединицы 2 цитохром-с-оксидазы, субъединицы 4 NADH-дегидрогеназы, субъединицы 6 ATP-синтетазы, цитохрома b и некоторых других. Увеличение транскрипционной активности указанных генов при снижении в трансформированной и опухолевой клетках количества митохондрий связывается с необходимостью поддержания в них нуж-ной скорости дыхания, с созданием и (или) поддержанием злокачественного фенотипа этих клеток. Здесь действительно просматривается ответная «компенсирующая» реакция клеток на ослабление в них окислительного фосфорилирования, но какие конкретно особенности или обстоятельства внутри опухолевых клеток индуцируют указанный адаптивный ответ, пока точно неизвестно.
Можно предполагать, что в митохондриях, как и в ядре, экспрессия некоторых генов рО2- или АФК-зависима. Известны, по крайней мере, несколько работ (Murphy et al., 1984; Suzuki H. et al., 1998), в которых рассматривается О2-зависимый механизм регуляции количества дыхательных ферментов. По этой логике, в опухолевой клетке с её относительной гипероксичностью (см. п. 2.1.3) экспрессия каких-то митохондриальных генов окислительного фосфорилирования и должна возрастать. Однако в тех же условиях опухолевой клетки эти адаптивно-защитные меры оказываются малоэффективными, так как синтезируемые ферменты дыхательной цепи частично подвергаются прямой или косвенной (через ПОЛ внутренней мембраны митохондрий) инактивации.
Есть ещё работа (Макаренко и др., 1992), в которой обнаружена исходно достоверная разница в содержании митохондриальных цитохромов в ткани печени между низко- и высокоустойчивыми к гипобарической гипоксии кры-сами. Как оказалось, у первых содержание цитохромов b, c, c1 и a было достоверно выше, чем у вторых. Связь степени устойчивости к гипоксии с количеством дыхательных ферментов представляется нам естественной, но проявляется она косвенно, через уровень внутриклеточного рО2. При более высоком содержании цитохромов утилизация О2 митохондриями должна происходить интенсивнее, соответственно снижается и рО2 в клетке. Если такая клетка к тому же оказывается в условиях гипобарической гипоксии, то устойчивость её к последней будет, очевидно, низкой по сравнению с клеткой, где указанного снижения рО2 при малом содержании цитохромов не происходит. Приведённая интерпретация рассматриваемого факта согласуется с зависимостью содержания дыхательных ферментов в клетке от уровня рО2, и наоборот. По той же причине опухолевые клетки с недостаточностью их дыхания и относительной гипероксией должны быть устойчивы к гипоксии.
Еще по теме 2.1. Митохондрии как первичный объект повреждения ииндукции пероксидативного состояния в клетке:
- 1.4.5. Внутриклеточная гипероксия как результат первичного процесса старения митохондрий
- Тема №1. Патологическая анатомия: объекты и методы исследования. Аутопсия. Патология клетки как интегративное понятие.
- 1.3. Митохондрии – наиболее уязвимое и основное«стартовое» звено в старении клетки
- Глава 8. Повреждение клетки
- Нелетальные повреждения генома клетки
- Общая этиология повреждения клетки
- Повреждение мембраны клетки
- Основные общие механизмы повреждения клетки
- 6. Повреждения грудной клетки.
- Механизмы токсического повреждения систем энергообеспечения клетки
- Определение экспрессии Толл-подобных рецепторов (TLRs) на клетках крови, клетках кожи и клетках эпителия слизистой зева.
- Первичные повреждения шейного отдела позвоночника
- Повреждение периферических нервов при инвазии нервных структур опухолевыми клетками