крайние типы адаптации к тропическому И ПОЛЯРНОМУ КЛИМАТУ
Климатические особенности Арктики и Антарктики с чрезвычайно низкой среднегодовой температурой, крайне продолжительной зимой и совсем коротким летом создают совершенно особые условия существования гомойотермных организмов.
Еще более ста лет назад были измерены температуры тела у полярных животных (песца, полярного зайца-беляка) при температурах окружающей среды около —50°С (Parry et Lyon, 1825). Эти исследования обнаружили, что постоянство температуры тела может поддерживаться при разнице внутренней температуры (тела) и среды, достигающей 80° С (см. табл. 13).Таблица 13
Температура тела полярных млекопитающих и температура среды
(по Hesse, 1924)
| Ви і | Температуря (°С) | Разница | |
| тела | Срезы | ||
| Песец (Alopex lago- pus) | 39,6 | —33,0 | 74,6 |
| Волк (Canis lupus) | 40,3 | -32,8 | 73,3 |
| Заяц-беляк (Lepus ti- tnidus) ................................ | 38,3 | —29,4 | 67,7 |
| Снежная куропатка (Lagopus tnutus) . . . | 43,3 | -37,3 | 0,6 |
Постоянство температуры тела у некоторых полярных видов даже в условиях сильных холодов (до —25° С) было обнаружено в работе с песцами и лисицами (Ольнянская и Слоним, 1947); при этом оказалось, что теплопродукция (газообмен) в условиях значительного охлаждения не повышается. Слабое повышение газообмена при охлаждении на фоне несколько снижающейся температуры тела было обнаружено у серой крысы (пасюка) в условиях Заполярья — Кольский полуостров (Руттенбург, 1950, 1953).
На рисунке 49 приведены данные изменений газообмена и температуры тела у некоторых обитателей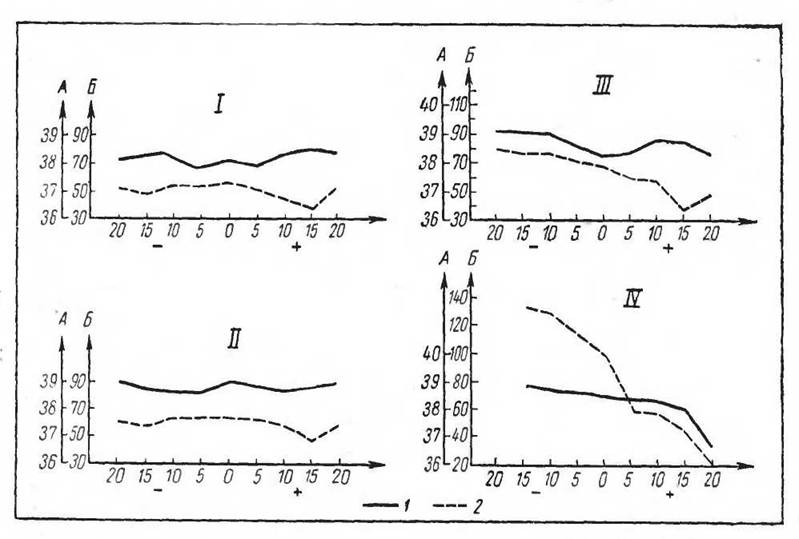
Рис. 49. Изменения газообмена и температуры тела при разных температурах среды у арктических млекопитающих (по Ольнянской и Слониму, 1947). / — песец белый; // — песец голубі й; /// — заяц-беляк; /V — заяц- русак:
а — температура тела (°С), б — потребление кислорода {мл)
Заполярья. Пределы выживаемости некоторых млекопитающих и птиц при низких температурах видны из табл. 14.
Таблица 14
Выживаемость некоторых животных при низких температурах (по Hart a. oth., 1957)
| Живо гные | Температура (°С) | Время выживания |
| Крупные млекопитающие Арктики | —40 | Неопре теленно |
| Кролик ....................................................... | —35 | 3,5—6,5 ч |
| Крупный рогатый скот.............................. | — 13 | 2 нетели |
| Крыса . | —35 | 4 л—120 мин |
| Утка................................. ......................... | — 10 | 384 и |
| Г олубь.................................................. | — 10 | 16—30 ч |
| Воробей . | -30 | 3—5 ч |
Продолжительность выживания при интенсивном охлаждении зависит главным образом от теплоизоляции, создаваемой шерстным и перьевым покровом животных. От этой теплоизоляции в значительной мере зависит и интенсивность повышения обмена веществ при охлаждении.
У арктических животных повышение обмена наблюдается только при очень низких температурах среды: у ласки, суслика и лемминга при —15—18° С и ниже; эскимосской собаки —24° С; песца —40° С; белого медведя —50° С (Scholander, Hock, Walters a. Irving, 1950). Исключительно постоянная температура тела наблюдалась как в состоянии сна, так и в периоды бодрствования у таких видов, как эскимосская собака, песец. При температуре среды —80° С температура тела оставалась постоянной на уровне 38° С. По этим же данным критическая точка у этих животных лежит ниже —50° С. Температура кожи у эскимосской собаки под мехом составляет около 30° С при температуре среды около —50° С. Однако температура носа, конечностей (например, у северного оленя) может не превышать в этих условиях 10° С (Irving, 1951). Вместе с тем при одинаковой теплоизоляции шерстного покрова устойчивость к охлаждению может быть различной. Так, при сравнении трех мелких млекопитающих Аляски — тундровой рыжей полевки (Clethrionotnys rutilus dawsoni), тундровой полевки (Microtus oeconomus macjarlani) и длиннохвостой землеройки (Sorex cincreus hollisteri), несмотря на одинаковую величину теплоизоляции, наиболее устойчивой к холоду оказалась рыжая полевка. Это особенно ярко было выражено при охлаждении в условиях движения воздуха. При этом характерными являются и позы животных при охлаждении, что также определяет устойчивость температуры тела (Morrison a. Tictz, 1957).Очень низкая температура критической точки обнаружена у горного козла (Creamnos americanus) на Аляске (Krogh a. Monson, 1954). При температурах среды от 20 до —20° С газообмен оставался постоянным, при
— 30° С наблюдалось повышение его на 23%. а при
— 50° С потребление кислорода повысилось на 30%. Температура — 20° С является средней для обитания этого животного.
На основании ряда исследований Шоландером предложена схема (рис. 50) изменений теплопродукции у тропических и арктических млекопитающих и птиц. Эта схема показывает, что у устойчивых к холоду гомойотермных организмов теплопродукция с понижением температуры среды нарастает очень медленно.
Это связывается Шоландером с высокой теплоизоляцией (см. выше). Тропические ф.ормы обладают большей реакцией на охлаждение. Поэтому они не могут длительное время существовать в условиях, когда температура снижается ниже— 10° С. Интенсивная химическая терморегуляция (по существу приближающаяся к величинам вершинного обмена — metabolisme de sommet) быстро приводит к истощению энергетических ресурсов и к гибели (Scholander a. oth, 1950).
У тропических видов — обезьян н хищных наблюдается ряд особенностей химической терморегуляции и ее взаимоотношений с физической, а также ряд особенностей теплоотдачи (Слоним и Щербакова, 1935; Слоним, 1937, 1941, 1952). Так, например, у представителей тропических и субтропических хищных —шакалов, динго — была обнаружена более высокая температура критической точки, свидетельствующая о том, что полное выключение химической терморегуляции наблюдается при более высокой температуре внешней среды. На рис. 37 приведены эти данные, полученные для домашней собаки, шакала и динго, содержавшихся на протяжении нескольких
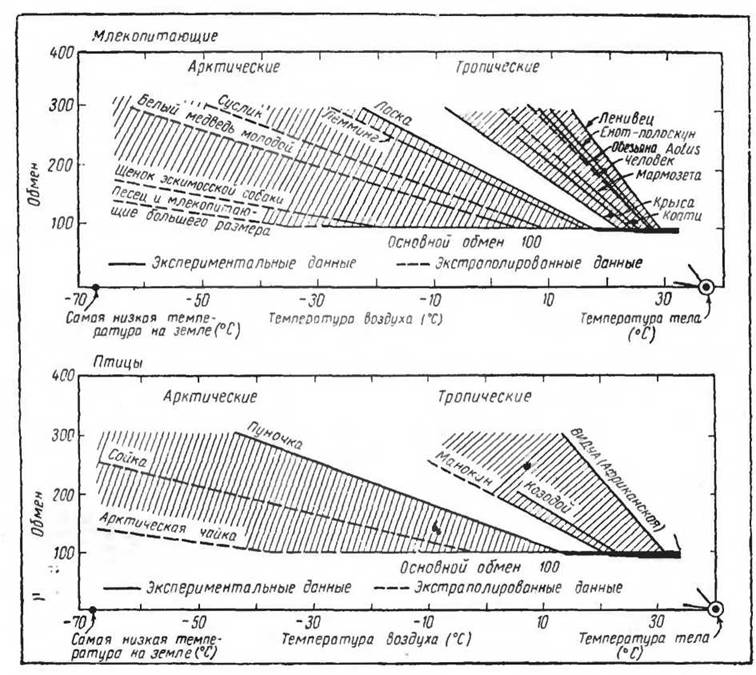
Рис. 50. Терморегуляция и термочувствительность арктических и тропических млекопитающих и птиц. Критические градиенты (термочувствительность) и наклон кривой зависят от произведения основного обмена и общей термоизоляции тела (по Scholander и сотр., 1950)
месяцев в одинаковых температурных условиях. На этих же объектах обнаружено, что интенсивность химической терморегуляции у тропических форм ниже, чем у обитателей умеренного климата. Например у шакала — обитателя субтропиков процент изменений обмена веществ при переходе от 10 к 25°С составляет около 4%, тогда как у собаки (с одинаковой примерно длиной шерсти) достигает 7%. То же имеет место и у тропической собаки динго.
Подобные же данные получены при сравнении обезьян— павианов и гамадрилов — обитателей плоскогорий Эфиопии (200 м н. у м.) и обезьян-макак (резус и ла- пундер).
У первых обнаружена очень интенсивная химическая терморегуляция (до 5% изменений обмена на ГС), а у макак значительно более слабо выраженная (до 2% на 1°С). Критическая точка была резче выражена у макак при более высокой температуре внешней среды.Следует отметить, что критическая точка лучше выражена у животных, обладающих механизмом полипноэ. В этом случае пониженная, по сравнению с низкими температурами, теплопродукция при дальнейшем повышении температуры среды или при увеличении длительности экспозиции сразу возрастает (Слоним, 1952). Так обстоит дело у всех хищных, жвачных (Brody, 1945), грызунов и насекомоядных (Веселкин, 1945; Слоним, 1952). У животных, обладающих потоотделением, например обезьян, лошади и у человека, критическая точка заменяется зоной постоянного уровня обмена, отражающего включение сначала сосудистой, а затем и потоотделительной реакции. На рисунке 51 приведены данные, полученные для разных видов обезьян. Проявление критической точки при разных температурах среды обусловлено тем, что непосредственно перед включением полипноэ обмен веществ более или менее длительное время оказывается пониженным. Это явление так называемой второй химической терморегуляции было описано (Kestner, 1924) и подробно изучено на разных видах (Слоним и Щербакова, 1938; Слоним, 1952). У тропических форм это понижение обмена при высоких температурах выражено лучше, чем у нетропических, что можно наблюдать у шакалов, динго, обезьян-макак. Вторая химическая терморегуляция, как правило, особенно хорошо выявляется при длительном умеренном нагревании, и это ограничение про-
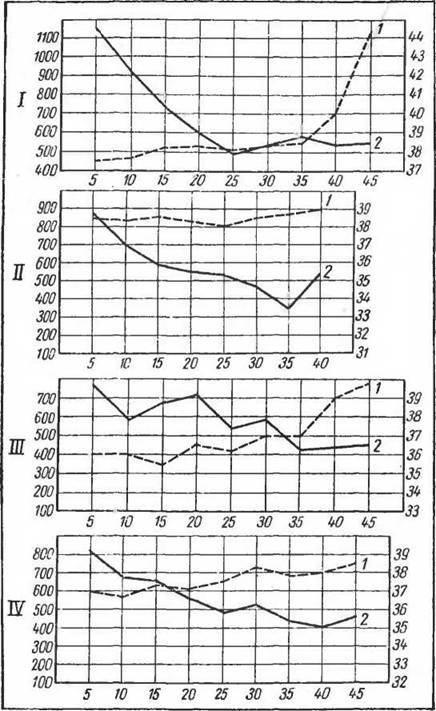
Рис. 51. Химическая терморегуляция и температура тела у низших обезьян (по Слониму и Щербаковой, 1940). / — павианы-гамадрилы; II — павианы-анубисы; III — макаки-резусы; IV — макаки-лапундеры. По оси ординат слева — О2 (мл!мин), справа — температура тела; по оси абсцисс — температура среды (°С):
1 — температура тела, 2 — потребление О2
дукции тепла, несомненно, имеет большое значение и в процессе осуществления животным мышечной деятельности.
Еще по теме крайние типы адаптации к тропическому И ПОЛЯРНОМУ КЛИМАТУ:
- Холодный климат
- Возбудитель тропической малярии (Plasmodium falciparum)
- Глава II. Полярные липиды грибов
- Возбудитель тропической малярии (Plasmodium falciparum)
- От антагонистической полярности к здоровой взаимозависимости
- 31. Психологический климат в коллективе
- Органосохранные операции при крайних проявлениях ППВР
- Жаркий климат пустынь
- Глава 4. Влияние концентрации фосфатов в среде на фракционный состав полярных липидов у других мицелиальных грибов
- Влияние веществ разной природы, полярности и концентрации на организацию молекул воды
- Клиническая картина крайне тяжелых форм острой лучевой болезни
- 45. ТЕРАПЕВТИЧЕСКИЙ КЛИМАТ.
- 27. Профессиональная адаптация. Причины трудностей профессиональной адаптации