Глава 4. Влияние концентрации фосфатов в среде на фракционный состав полярных липидов у других мицелиальных грибов
Представлялось интересным выявить характер влияния различного обеспечения фосфатами на синтез полярных липидов у некоторых других представителей мицелиальных грибов различного систематического положения.
В качестве объектов для исследования были выбраны следующие грибы: СлҐ coronafa, Enfv/rufenfQ, Л. tenuis 1120, Сип/. Рила fa 958,Л$/>. nigricans so и Pfen. chrgsogenum 63.
В таблице 8 суммированы некоторые данные, полученные при культивировании указанных выше грибов на синтетической среде №2 с различной исходной концентрацией фосфатов (0,1% -*+"Р и 0,0045% - - "-"Р вариант).
Из данных представленных в таблице видно, что при культивировании на синтетической среде с аспарагином (среда №2) исследованные грибы отличались по количеству накапливаемой биомассы.Однако, сравнительно высокое содержание биомассы (0,4 - 1,23 г сухой биомассы/ 100 мд среды) на относительно ранних этапах культивирования (48 часов) свидетельствует о том, что использованная синтетическая
- Ill -
среда благоприятна для развития большинства испытанных грибов (за исключением fnt viruienta , которая накапливала всего 0,25 г сухой биомассы/100 мл среды).
Таблица 8. Уровень биомассы и содержание в ней липидов у некоторых представителей мицелиальных грибов, выращенных на синтетических средах различного состава,
вариант среды
глюкоза! добавка (%) ! Р
культура гриба j (возраст 48 час)!
I
Ent. coronata
биом.абс. ісуммар- } поляр-
сух./Ю0мл}ные липи-{ ные липи среды }ды г/ІООгі ды (%)
’ j сух. биом.} нейтраль
5 і і ные липи
!! ды (%)
+ 0,4 31,9 8,4/91,6
0,4 29,8 4,5/95,5
Enf.
viru
tenta.
+ 0,25, 20,2 1,1/98,9
0,26 22,6 2,0/98,0
+
8
Alt 1
tenuis n +
Сиги. Рипліо. 8
A$p. 8 -
nigricans +
Pen. chry$o$enum 8
0,85 5,1 45,9/54,1
0,74 5,4 27,2/72,8
0,48' 7/9 26,3/73,7
0,58 8,6 28,9/71,1
0?5І 8/5 22,0/78,0
0,57 4,9 25,6/74,4
ї?23 6/2 26,1/73,9
0,83 4,6 14,4/85,6
0,66 8/2 24,8/75,2
0,51' 7,8 24,8/75,2
0,42 4?4 43,2/56,8
0,35 2,1 48,0/52,0
Содержание суммарных экстрагируемых липидов в мицелии различных грибов колебалось от 2,1-4,4 до 29,8-31,9%.
Количество липидов у всех представителей дейтеромицетов было невысоким и составляло от 2,1 (Pen. ehrg sogenu т ) до 8,6% (Alt. tenuis ), в то время как представители р. Entomopkihor^ синтезировали от 20 ( £.ni, vi- rulentcL) до 32% ( Int. coronata )липидов.Кроме того, были отмечены значительные колебания в процентном содержании полярных и нейтральных липидов у грибов различных таксонов. Дейтеромицеты характеризовались высоким содержанием полярных липидов, которые в отдельных случаях составляли почти половину ОТ суммы ЛИПИДОВ (напр. У tenu/S и Pen. cbrysogenum ), тогда как липиды энтомофтор были представлены почти исключительно нейтральными липидами.
Представлялось интересным изучить практически не исследованный вопрос о влиянии концентрации фосфатов на фракционный состав полярных липидов указанных выше грибов.
На рис.22а представлены результаты фракционирования полярных липидов £ni. coronata из которых следует, что кроме сфингомиелина (фракция I), ФЭА (фракция 6), ФХ (фракция 4), цереброзидов (фракция 8) и двух неидентифицированных липидов, в составе полярных липидов имеются фракции, которые по реакциям со специфическими реагентами являются бесфосфорными гликолипидами (фракции 2 и 3), обнаруживаются они только в мицелии "-”Р варианта.
Хроматографический анализ полярных липидов E„t. viruEenia выявил, что гликолипидные фракции присутствовали как в "-"Р, так и в "+"Р варианте, однако, если гликолипиды в "+"Р варианте синтезировались одновременно с фосфолипидами, то в "-”Р варианте они синтезировались на фоне резкого снижения фосфолипидов и являлись преобладающими фракциями в составе полярных липидов гриба (рис.226),
На рис. 22в?представлен состав фракций полярных липидов flti. {enuisу ввделенных из гриба после культивирования на средах с различной исходной концентрацией фосфора и глюкозы. Из рисунка видно, что и здесь присутствуют гликолипиды (фракции I и 2), обнаруживаемые в большом количестве в "-"Р варианте на среде с исходной концентрацией глюкозы 8%; фракция гликолипида №2 была также выявлена в *’+”Р варианте, однако, в меньших количествах.
Снижение- ття -
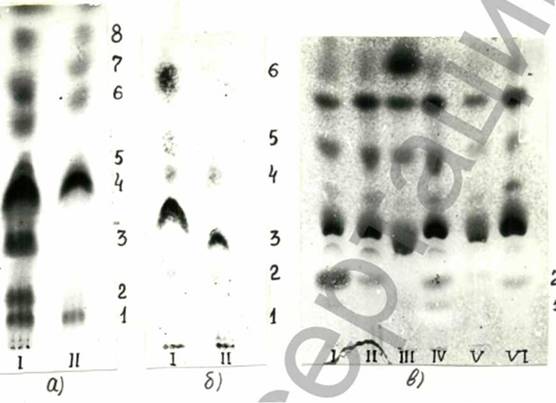
Рис.22 Фракционный состав полярных липидов,экстрагированных из мицелия £'nt coronota (a), £nt. virufenta (б),
/?£{. tenuis и Curv. Eunaio. (в):
а) I - "-"Р, II - "+"Р вариант.
Фракции: І-сфингомиелин, 2 и 3 гликолипиды, 4-ФХ, б- ФЭА, 8- цереброзид, 5 и 7 неидентифицированные фракции;
б) I - "+"Р, II - ”-"Р вариант.
фракции: 2 -гликолипид, 3 - ФХ, 6 - ФЭА;
в) І-ІУ - липиды Mt. tenuis, У,У1 - липиды Curv. funata, I - "-”Р, II - ”+"Р вариант (на среде с 8% глюкозы), III - "+’’Р, ІУ - ”-”Р вариант (с 2% глюкозы),
У - "+”Р, УІ - И-”Р вариант ( с 2% глюкозы).
Фракции I и 2 - гликолипиды.
исходной концентрации глюкозы в среде в 4 раза (т.е. до 2%) не препятствовало синтезу гликолипидов, однако, в этом случае 6ни были выявлены только в мицелии ”-”Р варианта. Очевидно, процессы липогенеза определяются в данном случае не только наличием фосфатов, но и другими компонентами питательной среды, например, сахаром.
Фракции гликолипидов были также идентифицированы при анализе полярных липидов, выделенных из мицелия другого дейтеромицета (Сиги lunato. ). В этом случае гликолипиды присутствовали в обоих вариантах опыта, однако в ”-"Р варианте отмечалось более интенсивное окрашивание пятна на ТСХ при реакции с антроновым и нафтоловым реагентами (на сахара).(рис.22в).
Аналогичные результаты были получены при изучении фракционного состава полярных липидов грибов fisp. nigricans и Pen. cJ>rysoger)um: гликолипид (фракция 2) присутствовал только в мицелии ”-"Р варианта, если гриб flip, nigricans культивировали на среде с 2% глюкозы, и фракция гликолипида обнаруживалась в мицелии грибов nigricans и Pen.chrysogewt/n обоих вариантов (”+” и ”-"Р) при использовании сред с исходной концентрацией глюкозы 8%.
В результате проведенных экспериментов установлено, что на используемой нами синтетической среде, все исследованные грибы синтезировали ранее не описанные у изучаемых организмов бесфосфорные гликолипиды.
При этом, дефицит фосфатов в среде усиливал накопление гликолипидов при одновременном снижении доли фосфолипидов в составе полярных липидов грибов.Таким образом, выявленная способность trapora к синтезу гликолипидов и стимуляция этого синтеза дефицитом фосфатов в среде, свойственна и другим мицелиальным грибам.
Полученные результаты можно рассматривать в свете гипотезы Роузера (Kowser ei. at., 1971),разделяемой советскими химиками
Л. Д. Бергельсоном и С.Г.Батраковым ( Бергельсон, 1984; SafraAou,
frerqetson, 1978) о возможной взаимозаменяемости мембранных липидов. Экспериментальное подтверждение эта гипотеза получила в работах с прокариотами. В частности, имеются сообщения Миникина с соавт о том, что при дефиците фосфатов происходит заїдена фосфолипидов на гликолипиды и пептидолипиды у некоторых видов бацилл и псевдомонад (tiinnihin et.at, 1971;1972;Х974). При недостатке фосфора
в среде, в составе полярных липидов Str.ottvQceus и других актиномицетов, была обнаружена липоаминокислота, представленная ор- нитинолипидом, которая более, чем на 2/3 (а иногда и полностью) заменяла фосфатидилэтаноламин в составе полярных липидов (Батраков с соавт.,1977; Касымбекова,Конова 1979; Конова,Касымбекова,1981).
В наших экспериментах впервые продемонстрирована возможность направленной модификации полярных липидов у представителей мицелиальных грибов различного систематического положения. Выявлено, что взаимозаменяемыми липидами являются фосфо- и гликолипиды.
РЛы полагаем, что механизм замены фосфолипидов бесфосфорными липидными компонентами в условиях, вызывающих снижение доли фосфолипидов (например, исходный дефицит фосфора в среде или исчерпание его запасов в процессе культивирования), является общебиологическим и действует на разных уровнях организации. Очевидно, механизм взаимозаменяемости мембранных липидов был выработан в процессе эволюции для сохранения жизнеспособности микроорганизмов в постоянно меняющихся условиях обитания. Физиологический смысл этого процесса, возможно, заключается в том, что бесфосфорные липиды мембран (такие, например, как гликолипиды) берут на себя роль фосфолипидов и подцерживают функцию мембран.
Кроме того, эти соединения могут включаться в ряд биохимических процессов (например, синтез полисахаридов), направленных на адаптацию организма к изменившимся условиям.- ІІб -
Анализ водно-метанольного слоя липидов некоторых грибов
С целью выявления возможного присутствия сильно полярных гли- косфинголипидов типа полигликозилцерамидов и ганглиозидов, проводили анализ водно-метанольного слоя, полученного в результате отмывки хлороформ-метанольного липидного экстракта грибов водой.
Липиды экстрагировали из биомассы грибов В?, trispora, Ent.co- ronata и £nt. viruferfta, выращенных в условиях достатка С*+”Р) и дефицита (”-"Р) фосфатов в среде.
После предварительного диализа материал анализировали методом ТСХ в системе растворителей хлороформ-метанол-вода (65:25:4).
В результате фракционирования было обнаружено две фракции (рис.23), которые давали положительную реакцию с антроновым и нафтоловым реагентами на сахар. Для установления липидной природы обнаруженных фракций была проверена полнота их очистки. С этой целью проводился повторный длительный диализ с последующим контролем методом: ТСХ. В результате было выявлено, что размер пятна фракции заметно уменьшался, что указывало на нелипидную природу этого соединения, присутствие которого могло быть связано с неполной очисткой содержимого диализного мешка от сахарных примесей. Вероятнее всего, фракция №2 имеет не липидную природу, а представлена исключительно сахаром.
Что хе касается фракции №1, то поскольку величина пятна на ТСХ не уменьшилась после повторного длительного диализа, можно думать, что эта сильно полярная фракция имеет липидную природу.
Как нам кажется, заслуживает внимания дальнейшее изучение обнаруженной у зигомицетов сильно полярной липидной фракции, т.к. до сих пор соединения подобного рода у грибов не были найдены, за исключением некоторых представителей Weurospora у которых был обнаружен тетрагексозилцерамид (Lester et.aP., 1974).
I.

Рис.23 Фракции водно-метанольного слоя липидов грибов &C.trispora.
(І,ІІ), мгопаїа. (Ш,1У) иt, гіги (vita. (У, у I);
I,III,У - "+"P, ІІ,ІУ,УІ - ”-"P вариант
1 IL Ш IV V VI
/

Глава 5. Состав жирных кислот yZ3P. irisporo, и других грибов в зависимости от содержания фосфатов в среде
Исследование липогенеза непременно включает изучение состава жирных кислот, т.к. они являются одним из основных компонентов, определяющих свойства липидов. Состав жирных кислот важен также для решения вопроса о практическом использовании липидов.
В связи с этим, изучали композицию жирных кислот в динамике развития hl tri tрога в зависимости от содержания фосфатов в сре
де. Исследования проводили на синтетических средах, обеспечивающих синтез жирных кислот только de novo
В таблице 9 представлены данные о составе и содержании отдельных жирных кислот, полученных из суммарных экстрагируемых липидов hl trispora после 96 часов культивирования гриба на средах с различной исходной концентрацией фосфатов (опыты по схеме "от спор"). Таблица 9. Состав и содержание жирных кислот в липидах BP. trispora
в зависимости от концентрации фосфатов в среде
| зкирные кислоты | содержание 0 | варианты опыта (% фосфатов) і отдельных жирных кислот (% от суммы) | |
| 0,05 | 0,5 | ||
| С14:0 | 6,20 | 2,3 | 4,67 |
| С14:1 | 1,80 | 1,7 | 1,48 |
| Сх | ІД4 | ||
| С16:0 | 40,91 | 34,35 | 27,24 |
| С16:1 | 2,82 | 2,47 | 3,34 |
| С18:0 | 7,78 | 10,27 | 9,24 |
| С18:1 | 17,35 | 19,35 | 16,89 |
| С18:2 | 21,94 | 29,53 | 37,08 |
| йодное число | 55,5 | 70,1 | 81,8 |
Из таблицы видно, что основными жирными кислотами, которые подвержены наиболее заметно?^ влиянию концентрации фосфатов, являются пальмитиновая (Cjg.g) и линолевая (Cj8;2) кислоты. Дефицит фосфора в среде способствовал преимущественному синтезу насыщенной кислоты, количество которой в этом случае составляло более 40% от суммы жирных кислот. Одновременно синтезировалось около 22% С18:2 кислоты. При исходной концентрации фосфатов в среде 0,05%, происходили изменения в соотношении этих жирных кислот: снижалось содержание пальмитиновой кислоты (до 34,35%) и повышался уровень линолевой кислоты (до 29,53%). При увеличении исходной концентрации фосфатов в среде до 0,5%, отмеченная тенденция становилась более явной, т.е. Cjg.Q кислота составляла 27,24%, a Cj8.2 кислота - - 37,08% от суммы жирных кислот.
Таким образом, было обнаружено, что недостаток фосфора в среде приводит к снижению ненасыщенности липидов за счет повышения отношения Ci6;(/Cjq:2. В данном случае это отношение составляло 1,86; 1,16 и 0,73 соответственно в ”0”, ”0,05” и "0,5”Р вариантах опыта.
Аналогичные результаты были получены при анализе жирных кислот нейтральной, преобладающей (более 70% от суммы липидов) фракции липидов Z3A tnsporo. 701 (+) при культивировании гриба в течение 96 часов на синтетической среде К?1 с различной исходной концентрацией КН2Р0^ (табл.10). В данных условиях эксперимента отношение ^16:0^^18:2 составляло 1»81 в условиях дефицита фосфора и соответственно 0,7 и 0,32 в ”0,5" и "0,05"Р вариантах. Более высокое значение отношения в случае *'0,5"Р варианта, по сравнению с ’'0,05"Р вариантом обусловлено тем, что увеличение исходной концентрации фосфатов до 0,5% вызывало в этом опыте (правда меньшее, чем в "бесфосфорном" варианте) повышение содержания Cjg.Q кислоты и снижение уровня Cjg.j И CjQ.j КИСЛОТ, при ЭТОМ, количество Cjq.2 кислоты не изменялось. При дефиците фосфатов было также отмечено некоторое снижение оодерасания и Cjg.j кислот, однако, количест
во Cjq.j кислоты при этом, оставалось на уровне, отмеченном в w0,05wP варианте.
Таким образом, была отмечена лабильность жирнокислотного компонента липидов в зависимости от обеспеченности клеток гриба фосфатами.
Таблица 10. Содержание жирных кислот в нейтральных липидах Ы trispora 701 (+) в зависимости от обеспеченности фосфатами
| жирные кислоты | варианты опыта (% КНоРО^, срела №1) содержание отдельных жирных кислот (% от суммы) 0 0.05 0,5 | ||
| С14:0 | 2,23 | 2,72 | 2,84 |
| С14:1 | следы | 0,02 | 0,76 |
| С16:0 | 40,98 | 13,13 | 27,7 |
| С16:1 | следы | 11,9 | 6,2 |
| С18:0 | 9,2 | 6,7 | 7,13 |
| С18:1 | 25,0 | 24,7 | 15,8 |
| С18:2 | 22,6 | 40,7 | 39,5 |
| йодное число | 60,8 | 102,4 | 87,7 |
В связи с тем, что фазы развития гриба могут быть сдвинуты во времени в зависимости от концентрации фосфатов, а также в связи с вероятностью изменения состава жирных кислот в процессе развития культуры, более четкие данные о составе жирных кислот в зависимости от концентрации фосфатов, могут быть получены при анализе жирных кислот в динамике развития культуры.
Результаты, полученные при анализе жирных кислот суммарных экстрагируемых липидов в процессе развития trizpora на трех вариантах синтетической среды, представлены в таблице II. Видно, что отмеченная выше тенденция .снижения содержания ненасыщенных жирных кислот при дефиците фосфора в среде проявлялась на протяжении всего процесса культивирования гриба.
Таблица II, Состав и содержание жирных кислот суммарных липидов Sf. trispora в процессе культивирования гриба на трех вариантах синтетической среды Р-3
жирные
кислоты
С12:0
Сх
С14:0
С16:0
С16:1
С18:0
СІ8:І
СІ8:2
С18:3
Сх
йодное
число
возраст культуры (час) и варианты опыта (% фосйатов)
содержание отдельных жирных кислот (% от суммы)
| инокуі ЛЯТ І і о | 41 час | 0 | ----------- р. 65 час ! t | ІІЗ час 0 0,05 0.5 | ||
| 0,05 | 0.5 | 0,05 | 0,5} | |||
| 0,7 J | 0,1 | Ы | 0,2 | І і • | ||
| 12,3 і | 1,5 | 0,3 | 1 і | |||
| 2,9 j 1,8 | 2,0 | 1,6 | 2,1 | 1,3 | І,Зі | 0,9 1,3 1,4 |
| 23,3 і35,3 | 23,8 | 26,9 | 30,2 | 24,8 | 25,9 j | 29,3 25,4 23;5 |
| 2,6 j 2,2 | 2,3 | 1,6 | 1,7 | 2,8 | I,7j | 1,8 2,5 2,4 |
| 4,3 |l0,7 | 8,1 | 6,3 | 7,4 | 8,5 | 6,7j | 6,3 6,5 5,8 |
| 16,6 |2I,i | 23,4 | 23,3 | 23,8 | 23,4 | 20,9 j | 22,8 17,6 17,7 |
| 29,9 }29,0 | 40,5 | 40,5 | 32,5 | 38,8 | 43,6} | 39,0 46,6 49,4 |
| 5,3 j | і і | |||||
| 2,1 | 1 | і і ( | |||||
| 82,3 j70,4 | 92,2 | 91,7 і | І 78,3 | 89,9 | 95,1 і | 88,8 98,8 І03,і |
Необходимо отметить, что в молодой культуре (41 час), уровень ^18:2 кислоты гораздо выше в ’’фосфорных” вариантах, но при этом не было разницы в процентном содержании этой кислоты в ”0,05” и ”0,5”Р вариантах. Однако, на более поздних стадиях развития, содержание линолевой кислоты тем выше, чем больше фосфора в среде. Динамика изменения отношения кислот Cjg.Q/Cjg.g в процессе развития tri- Spora представлена на рис.24, из которого видно, что наиболее высоким это отношение является при дефиците фосфора в среде и мало отличается при концентрации фосфатов 0,05 и 0,5%. Кроме того, была отмечена тенденция постепенного снижения этого отношения по мере развития культуры, что особенно отчетливо проявлялось в "бесфосфорном" варианте.Подобное явление происходило из-за повышения доли
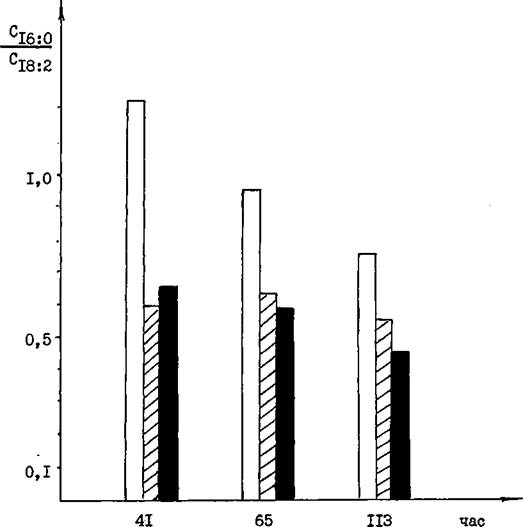
РПС.24 Изменение отношения жирных КИСЛОТ Cyg.g/CjQjg в процессе культивирования В/. trispora на средах с различной исходной концентрацией фосфатов: О - дефицит фосфатов; и - 0,05# фосфатов;
■ - 0,5# фосфатов.
линолевой кислоты на фоне постоянного содержания пальмитиновой кислоты. Указанное изменение в соотношении двух жирных кислот приводило к возрастанию ненасыщенности липидов.
Из представленных результатов очевидно, что концентрацию фосфатов в среде можно рассматривать как фактор, регулирующий ненасы- щенность липидов. Вероятнее всего, регуляторная роль фосфора осуществляется на уровне фосфолипидов. В настоящее время имеются убедительные доказательства о потребности отдельных фосфолипидов для активности десатуразных систем (baker, Lynen, 1971; Pugh,
Hales, 1973,1975; Ta (to. mo et. a?., 1973; Wat son, Rose», 1979;
Ga&rietides at. at., 1982).
В наших экспериментах было показано, что исключение фосфатов из опытной среды приводит к значительному снижению доли фосфолипидов в составе полярных липидов. Одновременно при этом был отмечен усиленный синтез бесфосфорных компонентов в составе полярных липидов, среди которых преобладали гликолипиды.Отмеченное снижение основных фосфолипидных компонентов сопровождалось повышением насыщенности липидов. Полученные результаты позволяют считать, что для процесса десатурации, обеспечивающего синтез С^д.£ кислоты у
trispora. требуется определенный уровень фосфолипидов в клет
ках.
Однако, необходимо учитывать, что проявление указанной зависимости во многом определяется физиологией культуры в каадом конкретном случае. Так, при культивировании ЬР. trispora на "+" и ”-”Р вариантах синтетической среды №2 с исходной концентрацией глюкозы 8%, нам не удалось выявить значительного повышения насыщенности при недостатке фосфора в среде. В данных условиях эксперимента, содержание липидов в клетках практически не менялось в процессе культивирования, а оставалось на уровне посевного материала. При этом было отмечено лишь очень незначительное повышение отношения Cjg.Q/Cjg.g в условиях дефицита фосфатов. Отношение указанных кислот в ”+”Р варианте составляло 0,75; 0,74; 0,68 и в ”-"Р варианте - 0,83; 0,83 и 0,66 после 48, 96 и 144 часов роста гриба соответственно. Относительная стабильность состава жирных кислот, отмеченная в указанном опыте, мояет быть связана с отсутствием активного синтеза липидов в рассматриваемых условиях эксперимента.
Таким образом, соотношения отдельных жирных кислот могут претерпевать большие или меньшие изменения в зависимости от физиологического состояния культуры, связанного с составо?.! используемых сред.
Следующим этапом наших исследований явилось изучение влияния концентрации фосфора в среде на синтез жирных кислот другими ми- целиальными грибами, описанными выше.
Исследования проводились на синтетических средах, отличающихся также исходной концентрацией глюкозы. Все анализы проводились через 48 часов культивирования грибов. Полученные результаты представлены в таблицах 12 и 13.
Для всех исследованных грибов, у которых в составе жирных кислот преобладают Cjg.£ и Cjg.Q кислоты, было установлено большее ИЛИ меньшее увеличение отношения Cjg.Q/Cjg.g.npH дефиците фосфатов в среде. В случае Сиги Zunata , у которой преобладает С18:1 кислота, эти отношения совпадали в ”+” и ”-”Р вариантах (табл.12).
Испытанные представители энтомофтор отличались по типу синтезируемых липидов: Ent. coronet а преимущественно синтезирует насыщенные липиды с преобладанием в их составе миристиновой (Cj^.q) кислоты (45,2-46,5% от суммы кислот). У второго энтомофильного гриба преобладают Cjg.| и Cjg.j кислоты. В обоих случаях практически не отмечалось влияния фосфатов на соотношение насыщенных и ненасыщенных жирных кислот (табл.13).
Таблица 12. Состав жирных кислот некоторых представителей дейтеромицетов, культивируемых на различных синтетических средах
куль
тура
гриба
варианты
среды
ГЛЮ-!
С12:0
жирные кислоты (% от суммы) CI4:0 С16:0 С16:1 С18:0 С18:1 С18:2
С18:3
16:0
18:2
flit, tenuis
+ 4,94 31,18
- 1,82 3,94 30,71
3,51 30,51 29,84 4,57 32,23 26,72
!I,04
і 1,15
8,86
6,53 21,14 2,25 33,44
2,44 25,44 35,57 3,41 31,38 29,52
Ю,6
Сиг\/.
ianata
0,72 27,74 5,21 3,65 36,93 25,75 1,4 29,94 5,15 3,01 31,64 28,84
1,08
1,05
nigricans
+ 0,21 0,99 15,8 1,56 5,52 26,34 45,31 4,25 - 0,51 1,8 21,94 2,11 6,2 30,7 34,74 2,01
0,66 16,76 следа 4,7 28,44 45,01 4,41 0,64 20,32 4,68 28,9 42,2 3,26
0,35
0,63
0,37
0,48
Pen. dvtj Sogenum
1,18 15,02 4,37 3,91 23,6 46,9 5,0 3,9 22,2 следа 8,0 24,3 37,9 3,71
0,32
0,6
Таблица ІЗ, Состав жирных кислот двух представителей энтомофтор, культивируемых на средах с различной концентрацией фосфатов
| гриба ! щ ! | р | !С12:0 | С14:0 | с16:0 | С16:1 | С18:0 | С18:1 | С18:2 С20:4 |
| Ent. | + | 12,6 | 46,5 | 16,1 | - | 2,0 | 12,9 | 10,0 |
| Coronata. | - | 12,7 | 45,2 | 16,5 | - | 1,9 | 15,1 | 8,8 |
| £nt. | + | 2,5 | 15,1 | 22,8 | 16,7 | 2,9 | 30,0 | 4,2 6,0 |
| virulenta | о» | 0,6 | 8.9 | 21,1 | 20,0 | 3,1 | 32,4 | 5,5 6.5 |
| Было отмечено, что | ПОЧТИ | во всех случаях, ' | снижение Cjg.p кис- | |||||
лоты при недостатке фосфора коррелировало с некоторым повышением уровня олеиновой (Cjg.j) кислоты. Это, очевидно, связано с тем, что при снижении десатуразной активности, ответственной за синтез линолевой кислоты, непосредственный субстрат реакции, олеиновая кислота накапливается в несколько больших количествах.
Были выявлены некоторые особенности в жирнокислотном составе
в зависимости от исходной концентрации глюкозы в среде. Если гриб ДР1. tenui s выращивался на среде с 8% глюкозы, то отношение
^16:0^^18:2 ПРИ ДеФиЧите фосфора в среде равнялось 1,15, что лишь немного превышало соответствующее значение для ”+"Р варианта, в котором оно составляло 1,04. При исходной концентрации глюкозы 2%, наблюдалась более четкая разница: отношение составляло соответственно 0,6 и 1,1 в '*+*' и "-“Р вариантах опыта.
В добавок к этому, при сравнении ”+"Р вариантов с 8 и 2% глюкозы в среде, выявляются следующие особенности: приболев низкой концентрации глюкозы, на фоне одной и той же питательной среды, было отмечено более высокое содержание короткоцепочечных насыщенных жирных кислот (Cj2:o и C^.q), которые составляли в сумме 15,39%, тогда как на среде с 8% глюкозы было выявлено всего 4,94% (от суммы жирных кислот) миристиновой (Cj^.q) кислоты. Прослеживалась также заметная разница в содержании преобладающих жирных кислот: Cjq.q, Cj£.j и Cjq;2* При снижении исходной концентрации в среде в 4 раза (с 8 до 2%) в составе жирных кислот снижалась доля ^16:0 и ^18:1 кислот ПРИ одновременном повышении уровня Cjg^ кислоты. Таким образом, у Att. tenuis синтез линолевой кислоты протекает более интенсивно при низких концентрациях глюкозы в среде. Исключение фосфатов ив опытных сред в обоих случаях (на 8 и 2% глюкозы) приводило к увеличению отношения Cjg.Q/Cjg.2, однако, как указывалось выше, наиболее отчетливо эта тенденция проявлялась на среде с 2% глюкозы.(табл,12).
Совершенно иная картина наблюдалась для А/>./7/^г/слл?.Во-1х, у этого гриба не было отмечено каких-либо существенных отличий в содержании пальмитиновой, олеиновой и линолевой кислот в "+"Р вариантах опыта, отличающихся исходной концентрацией глюкозы в среде (табл.12). Создание условий дефицита фосфатов, в отличие от Ati. tenuis приводило к более значительному смещению отношения
Cl6:(/Ci8:2 в повышения на среде с 8% глюкозы. В этом слу
чае были получены значения 0,63 и 0,35 (по сравнению с 0,48 и 0,37 на 2% глюкозы) в и "+"Р вариантах опыта соответственно.
Таким образом, обобщая представленные результаты, можно сделать следующие выводы: у большинства проанализированных грибов, как и у irispora выявлена зависимость синтеза линолевой ^18:2^ кислоты от концентрации фосфатов в среде. Дефицит фосфора приводит к увеличению насыщенности липидов, в основном, за счет повышения отношения Cjg.Q/Cjg.g. Отмеченная тенденция может проявляться в различной степени в зависимости от индивидуальных особенностей гриба и физиологических условий культивирования. В частности, установлено, что одним из существенных факторов, влияющих на соотношение отдельных жирных кислот и регуляторную роль фосфора, является концентрация глюкозы в среде.
В связи с вышесказанным важно отметить, что для ряда микроорганизмов показано значение повышения насыщенности связей жирных кислот к устойчивости клеток к неблагоприятным условиям внешней среды (Васюренко,Синяк,1976;Синяк,рудиченко,I960). Возможно в этом и заключается физиологический смысл обнаруженного нами явления увеличения насыщенности липидов некоторых мицелиальных грибов при недостатке фосфора в среде.
Во всех случаях повышение насыщенности липидов коррелировало с изменениями во фракционном составе полярных липидов, а именно со снижением доли фосфолипидов в их составе. Как уже отмечалось выше, можно полагать, что процесс синтеза Cjg.g кислоты у изученных нами мицелиальных грибов связан с определенным составом и содержанием полярных липидов в клетке.
Раздел II. Изучение возможного практического использования липидов грибов
Известно, что растительные масла широко используются не только в пищевой промышленности, но и в других отраслях народного хозяйства, в том числе, в металлургии, лако-красочной, шинной промышленности. В связи с этим, актуальной является проблема замены пищевых растительных масел возможными их аналогами. В качестве таких аналогов испытываются липиды микробногсг происхождения. При этом, как указывалось выше (см.главу I лит.обзора), применение их может быть самым разнообразным.
Гриб /if trispora. являющийся продуцентом р -каротина к липидов уже нашел применение как источник кормовых препаратов. Известно его использование и для получения пищевых красителей, например, ликопина ( Скрябин с соавт.,1966;Бехтерева,1973; fineJerson,
1957; Js&r «
Еще по теме Глава 4. Влияние концентрации фосфатов в среде на фракционный состав полярных липидов у других мицелиальных грибов:
- Глава 2. Состав и содержание липидов 31 ігіірогл в зависимости от концентрации фосфатов в среде
- Глава II. Полярные липиды грибов
- Влияние веществ разной природы, полярности и концентрации на организацию молекул воды
- Фракционный состав нефти
- Химический и фракционный состав нефти
- Влияние острого введения алкоголя на предвидение и формирование пространственной памяти в виртуальной среде
- 2.3.2. Образующиеся в плазматической и других мембранах перекиси липидов в норме выполняют ряд необходимых и важных для клетки функций
- Влияние витаминов и других факторов роста
- Влияние ГУМТ на концентрацию глюкозы и С-пептида в пуповинной крови
- Влияние Ка20880-инозина на процессы перекисного окисления липидов у больных раком орофарингеальной области, получавших химиолучевую терапию