ПРИЛОЖЕНИЕ 2 Базовые методы генной инженерии
Полимеразная цепная реакция (ПЦР)
Амплификацию фрагментов ДНК проводят с помощью полимеразной цепной реакции. Принцип метода заключается в энзиматическом синтезе определенных фрагментов ДНК, фланкированных последовательностями нуклеотидов,
комплементарными последовательностям праймеров.
Праймеры - это короткие искусственно синтезированные олигонуклеотиды, являющиеся затравками для синтеза ДНК термостабильной ДНК- полимеразой.В настоящее время для решения разных задач используются разные ДНК-полимеразы. Главными качествами, по которым отличаются полимеразы для ПЦР, являются их скорость и точность синтеза ДНК. Как правило, эти два параметра взаимоисключающие - высокопроцессивные ДНК-полимеразы обладают невысокой точностью, а точные (proof-reading) полимеразы синтезируют ДНК медленнее, и, как следствие, не могут быть использованны для амплификации длинных фрагментов. Точность полимеразы определяется наличием у нее корректирующей 3'-5'-экзонуклеазной активности. При встраивании в синтезируемую цепь ДНК некомплементарного матрице нуклеотида такая полимераза способна заменит его на «правильный» и продолжить синтез. Обычные полимеразы в таких случаях либо прекращают синтез ДНК, либо продолжают его, оставив в составе цепи некомплементарный матрице нуклеотид (mismatch). Для решения большинства (но не всех!) молеклярно- биологических задач оптимально использование смеси точных и высокопроцессивных полимераз.
В состав 10-кратного буфера для ПЦР входят, как правило, следующие компоненты: 100 мМ Трис-HCl (pH 9,0); 500 мМ КС1, 1% тритон X-100. Необходимый компонент буфера - ионы Mg2'. Его концентрацию в составе ПЦР-смеси можно варьировать, как и температуру отжига праймеров, добиваясь получения гомогенного продукта. Реакция проводится в пробирках типа Эппендорф объемом 0,2 - 0,5 мл, предпочтительно с утоньшенными стенками, либо в плашках.
В качестве примера, можно использовать следующий состав реакционной смеси для ПЦР:
10х ПЦР буфер 2,5 мкл;
1 мкл;
1 мкл;
2 мкл;
10 мкМ «прямой» праймер (10-15 пмоль)
10 мкМ «обратный» праймер (10-15 пмоль)
10х dNTP 2,5 мкл;
25 мМ MgCl2 (если буфер не содержит Mg2+)
Taq-полимераза 0,5-1,5 ед;
ДНК 0,01-0,02 мкг;
MQ вода до 25 мкл.
2 мМ каждого
Внимание! Состав 10х dNTP: по дезоксирибонуклеозидтрифосфата.
праймера можно
Для примерного расчета количества пользоваться формулой: 1 пмоль 20-членного праймера составляет 0,0066 мкг.
Для предотвращения испарения на реакционную смесь наслаивают стерильное вазелиновое или минеральное масло (в большинстве современных амплификаторов предусмотрена возможность подогрева крышки пробирок, что делает добавление масла необязательным).
Простейший расчет температуры отжига праймера размером до 24 п.н. производится по следующей формуле:
Т=2 х п(А/Т) +4 х m(G/C) + (0.. .5),
где n, m - число соответствующих нуклеотидов, комплементарных матрице; Т - температура, °С. Оптимальными являются праймеры с температурой отжига 55-68°С. Время отжига праймеров варьирует от 5 до 30 c в зависимости от амплификатора.
Для большинства коммерческих ДНК-полимераз оптимальная рабочая температура равна 72°С. Время синтеза (элонгации) определяют в зависимости от длины амплифицируемого фрагмента (скорость работы Таq полимеразы примерно 1000 п.н./45 с).
Конструирование праймеров для ПЦР
Основной задачей при конструировании праймеров является достижение их специфичности к амплифицируемому участку ДНК. Эта задача решается введением достаточно длинной нуклеотидной последовательности, которая может быть достигнута только большим количеством совпадающих остатков. Чем длиннее олигонуклеотид, тем выше специфичность. Однако при длине праймера 25 нуклеотидов, гомологичного к уникальной последовательности, практически всегда можно добиться получения гомогенного продукта. Желательно, чтобы 3'-конец синтезируемого праймера точно совпадал с последовательностью ДНК-матрицы и содержал не более двух G или C на последние пять нуклеотидов.
При дизайне праймеров также следует учитывать соотношение между GC и AT нуклеотидами. Для АТ-богатых последовательностей праймер может получиться
низкоспецифичным и иметь низкую температуру отжига. Желательно, чтобы температура отжига была одинакова в прямом и обратном праймерах. В случае, если температура отжига праймеров разная, в ПЦР следует использовать наименьшую. Следует избегать наличия участков последовательности, способных сформировать высокостабильные вторичные структуры. Следите, чтобы в последовательностях прямого и обратного праймеров не оказалось комплементарных участков, которые могут обеспечить формирование стабильных дуплексов между праймерами. Для контроля отсутствия подобных дуплексов ставится контрольная реакция в которую вносятся все компоненты, кроме матричной ДНК. Для дизайнирования праймеров рекомендуется использовать специальное программное обеспечение (Primer3, Seqman).
Для облегчения клонирования в последовательность праймера вводят сайт узнавания какой-либо рестриктазы. Однако необходимо, чтобы с 5'-конца было несколько дополнительных нуклеотидов, что обеспечит успешную посадку фермента. Существуют таблицы, в которых определена минимальная длина выступающего фрагмента для наиболее часто используемых рестриктаз. Обычно длина этого дополнительного участка составляет 2-5 нуклеотидов.
При конструировании праймера для задачи направленного мутагенеза, необходимо, чтобы миспозиция приходилась не на конец праймера, а на его середину (так чтобы 3'-конец оставался полностью комплементарен исследуемому району).
Гель-электрофорез нуклеиновых кислот
Обычно электрофорез ДНК проводят в гелях агарозы. Области эффективного разделения линейных молекул ДНК в тысячах пар нуклеотидов в зависимости от концентрации агарозы в геле приведены в табл. П.2.1.
Электрофорез проводят в одном из двух буферов: трисацетатном (1хТАЕ) или трис-боратном (0,5хТВЕ) (состав буферов приведен в приложении). Для приготовления блока геля внесите соответствующую навеску агарозы в колбу, залейте свежеприготовленным буфером (удобно иметь 10-кратный запасной буфер и перед использованием развести его до однократного) и растворяйте агарозу при нагревании (нагревание лучше проводить в кипящей водяной бане во избежание пригорания агарозы или плавить агарозу в СВЧ-печи).
Таблица П.2.1. Области эффективного разделения линейных молекул ДНК в зависимости от концентрации агарозы в геле
| Концентрация агарозы, % | Размеры фрагментов, т.п.н. |
| 0,6 | 20-10 |
| 0,8 | 9-0,7 |
| 1,0 | 7-0,5 |
| 1,5 | 4-0,2 |
| 2,0 | 3-0,1 |
Для большинства экспериментов наиболее удобен горизонтальный вариант электрофореза. Перед заливкой геля установите заливочную площадку прибора строго горизонтально. После полного растворения агарозы остудите раствор до 50°С. Вставьте в пазы прибора ограничивающие пластинки. Залейте теплый раствор агарозы так, чтобы толщина слоя агарозы составляла 4-5 мм, предварительно вставив у одного из концов геля гребенку для формирования стартовых лунок. Гребенка не должна упираться в дно поверхности, на которой формируется блок геля. Зазор между дном лунки и основанием геля должен составлять 0,5-1,0 мм. После полного затвердения геля (30-40 мин при комнатной температуре, значительно быстрее на +4°С) удалите ограничивающие пластинки и залейте электродный буфер так, чтобы гель был залит слоем буфера на 0,5-1,0 мм. Аккуратно удалите гребенку, стараясь не повредить лунки. Подсоедините клеммы прибора к источнику питания (движение образцов ДНК идет в направлении от катода к аноду, т.е. от «-» к «+»).
Смешайте образцы ДНК, подлежащие разделению, с 1-2 мкл красителя (смесь 0,25% бромфенолового синего или ксиленцианола, 30% глицерина и 25 мМ ЭДТА) и с помощью автоматической пипетки внесите их в лунки геля. Наслаивание образцов проводите под буфер. Если необходимо определить размеры разделяемых фрагментов, в отдельные лунки внесите маркерные образцы (размеры маркерных фрагментов даны в приложении). Включите напряжение.
Целесообразно несколько минут давать малое напряжение (это позволяет сконцентрировать разделяемый материал на старте), после чего установить напряжение, необходимое для проведения разделения. Ход электрофореза контролируют по движению красителей. Для 1% геля подвижность бромфенолового синего соответствует фрагменту размером 300-500 п.н., а ксиленцианола - 3-4 т.п.н. ( в 6-8% полиакриламидном геле 40-50 п.н. и 150-170 п.н., соответственно). По окончании электрофореза отключите прибор от сети. Если гель исходно не содержал бромистого этидия, перенесите гель в ванночку и залейте раствором бромистого этидия (1 мкг/мл) и оставьте на 15 мин в темном месте.Бромистый этидий (3,8-диамино-6-этил-5-фенилфенатридиум бромид)
Синтезирован в 1950 г. как антитрипаносомный агент. Это соединение оказалось эффективнее в 10-50 раз. Его до сих пор используют при лечении трипанозомозов животных в субтропической и тропической зонах. Бромистый этидий интеркалирует (встраивается) в молекулы ДНК и образует ван-дер- ваальсовы взаимодействия с азотистыми основаниями. При этом изменяются параметры спирали - она удлиняется на 27%. Молекула ДНК поглощает на 254 нм, поглощение собственно бромистого этидия - при 366 нм, комплекс «нуклеиновые кислоты - бромистый этидий» флюоресцирует в области 590 нм (красно-оранжевая область спектра). Большая часть трансиллюминаторов дает свечение 254 или 302 нм (зависит от типа фильтра). Реакция между ДНК и бромистым этидием обратима, но диссоциация комплекса крайне медленна и измеряется днями. Для ускорения диссоциации этого комплекса рекомендуют провести его через катионобменную смолу Dowex- AG50W-X8, или экстрагировать ДНК органическим растворителем, например, изопропанолом или н-бутанолом.
Бромистый этидий эффективно связывается с РНК (по участкам, образующим шпильки) и суперскрученной ДНК. При этом молекулы бромистого этидия интеркалируют между соседними основаниями в ДНК. При последующем облучении ультрафиолетом образовавшийся комплекс дает свечение в краснооранжевой области спектра.
Часто для прямого визуального наблюдения хода разделения фрагментов электрофорез проводят в присутствии бромистого этидия. Для этого при приготовлении геля в охлажденную до 500С агарозу добавьте концентрированный раствор бромистого этидия до конечной концентрации 0,5 мкг/мл. Электрофорезный буфер в этом случае также должен содержать 0,5 мкг/мл бромистого этидия.Внимание! Обращаться с бромистым этидием необходимо осторожно, избегая его попадания на открытые участки тела, так как он является канцерогеном.
Концентрированный раствор бромистого этидия (3-10 мг/мл) хранится при 40С в темной склянке.
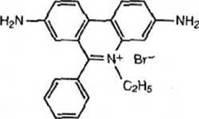
Рис. П.2.1. Молекула бромистого этидия
По окончании окрашивания гель ополаскивают холодной водой, переносят на полиэтиленовую пленку и просматривают под источником УФ-излучения.
Внимание! Помните, что при работе с источником ультрафиолета обязательно нужно пользоваться защитными очками.
Фотографирование гелей проводят в проходящем или отраженном УФ свете. Съемку проводят с использованием красного или оранжевого светофильтра. Можно использовать цифровые камеры. Если электрофорез проводился в присутствии бромистого этидия, часто не требуется дополнительного прокрашивания геля, достаточно просмотреть гель под источником УФ. Обычно для хорошего разделения фрагментов ДНК в лунку вносят такое количество ДНК, чтобы в индивидуальной полосе было не более 0,2 мкг ДНК. При наличии мощного источника УФ можно увидеть полосы, содержащие 0,02-0,05 мкг ДНК.
Обработка ДНК рестрикционными эндонуклеазами
Рестрикционные эндодезоксирибонуклеазы (рестриктазы) II класса, широко используемые в исследовательской практике, были открыты в 1970 г. Эти ферменты обладают способностью «разрезать» в определенных участках двойную спираль ДНК, разрывая сахарофосфатные связи в обеих нитях (для рестриктаз I и III классов место разрыва не однозначно связано с положением участка узнавания). Очевидная ценность ферментов II класса для генно-инженерных исследований инициировала их активный поиск в различных организмах, что к 2000 г. привело к обнаружению и очистке более 500 рестриктаз, из которых в научноисследовательской практике используется около 300.
Рестриктазы II класса обладают сравнительно малой молекулярной массой и для проявления активности требуют только ионов Mg. Эти ферменты узнают короткие, содержащие 4-8 звеньев, последовательности нуклеотидов, и большинство из них специфично расщепляет ДНК в пределах узнаваемого участка. Участки, узнаваемые этими ферментами, как правило, характеризуются наличием вращательной симметрии второго порядка, т.е. являются палиндромами. Для примера ниже приведены последовательности, узнаваемые часто применяемыми рестриктазами, и указаны места расщепления. Обычно указывается одна цепь этой последовательности в направлении от 5'- к 3'- концу. Примеры рестриктаз, часто применяемых в исследованиях, приведены в табл. П.2.2.
Таблица П.2.2
| Рестриктаза | Узнаваемая последовательность |
| EcoRI | g/аатгс |
| BamHI | G/GATCC |
| HindIII | A/AGCTT |
| SalGI | G/TCGAT |
| BglII | A/GATCT |
| SmaI | CCC/GGG |
| EcoRII | C/CAGG C/CTGG |
| AluI | AG/CT |
| Sau3AI | /GATC |
| BspRI | GG/CC |
| MspI | C/CGG |
| Cfrl3I | G/GNCC |
Рестриктазы могут разрезать ДНК в одном месте или места разрыва в двух нитях могут быть сдвинуты друг относительно друга в пределах сайта узнавания, таким образом ДНК, расщепленные рестриктазами, могут образовывать "тупые" или выступающие ("липкие") концы. В качестве примера показаны участки ДНК после расщепления рестриктазами BamHI и SmaI:
5'- NNG GATCCNN - 3' "липкие"
3'- NNCCTAG GNN - 5'
5'- NNCCC GGGNN - 3' "тупые"
3'- NNGGG CCCNN - 5'
За единицу активности рестриктазы принимается такое количество фермента, которое обеспечивает полное расщепление 1 мкг маркерной ДНК в течение 1 ч при 37°С.
Следует обязательно иметь в виду, что при определенных условиях может происходить нарушение строгой специфичности расщепления рядом рестриктаз. Это может наблюдаться для ферментов EcoRI, BamHI, HindIII и некоторых других. Вызывать подобные нарушения могут следующие причины:
неоптимальная ионная сила;
наличие таких органических компонентов, как этанол и глицерин;
замена Mg2+ на Mn2+;
высокая (избыточная) концентрация фермента.
Все вышесказанное предъявляет ряд обязательных требований к постановке реакции расщепления ДНК рестриктазами:
препарат ДНК должен быть хорошо очищен;
объем вносимого фермента не должен превышать 1/10
объема инкубационной смеси;
все растворы готовят на бидистилляте;
необходимо использовать оптимальный буфер для каждой рестриктазы.
Внимание! Препараты рестриктаз необходимо хранить при
температуре не выше -15°С и не ниже -20°С. В процессе работы фермент постоянно должен находиться в ледяной бане. Каждый отбор аликвоты рестриктазы проводите только новым наконечником.
Реакцию удобно проводить в микропробирках объемом 0,51,5 мл. Для аналитического варианта необходимо взять в пробу 0,5-1 мкг ДНК (в зависимости от числа ожидаемых фрагментов), добавить 1,5-2 мкл соответствующего концентрированного буфера, 2-3 ед. фермента и водой довести объем смеси до 1520 мкл. После внесения всех компонентов пробирку нужно закрыть крышкой, интенсивно встряхнуть и инкубировать в термостате при 37°С в течение 1-2 ч. Для некоторых рестриктаз оптимальная температура отличается от 37°С, в этих случаях смотрите паспорт фермента. Для остановки реакции чаще всего достаточно прогреть инкубационную смесь 15 мин при 65°С и затем охладить (в каждом конкретном случае следует проверить возможность инактивации на 65°С у конкретного производителя).
Реакция лигирования
Для образования ковалентной связи между молекулой векторной ДНК и исследуемым фрагментом ДНК проводят обработку смеси, содержащей оба вида молекул, ферментом ДНК- лигазой. Фермент образует фосфодиэфирную связь между З'-ОН и 5'-фосфатной группами концов ДНК.
Для проведения реакции векторную молекулу и образец исследуемой ДНК, обработанные соответствующими рестриктазами, смешайте в объеме 0,5-1,5 мл. Целесообразно, чтобы суммарное количество ДНК в смеси составляло 0,5-1 мкг. Молярное соотношение вектор/встраиваемая ДНК (точнее, соотношение концов линеаризованных молекул) желательно иметь 1/10. Это позволяет уменьшить вероятность обратного замыкания векторной молекулы самой на себя. Еще более эффективным способом уменьшения количества "пустышек" является дефосфорилирование векторной молекулы ДНК.
К смеси лигируемых ДНК добавьте 10х буфер для лигирования и рАТФ до концентрации 50мМ (многие буфера исходно содержат рАТФ).
Внесите в пробирку со смесью 0,1-1,0 мкл лигазы. Если лигирование проводится по "липким" концам, обогащенным GC парами, реакцию следует проводить при 12оС в течение 16-24 ч. В случае обогащения выступающих концов AT парами - при 5°С в течение 36-48 ч. Если лигирование проводится по "тупым" концам, реакцию проводят при 12°С в течение 36-48 ч, но концентрация лигируемых ДНК (как векторной, так и встраиваемой) должна быть на порядок выше, чем в предыдущем случае.
Солевая трансформация клеток Escherichia coli
Данная процедура предназначена для получения множества копий индивидуальных рекомбинантных молекул ДНК. Наиболее часто используемый вариант - трансформация клеток, обработанных хлоридом кальция. Обработка клеток СаС1 2 способствует более эффективному проникновению молекул ДНК из среды в клетки. Как правило, частота трансформации кольцевыми молекулами на несколько порядков выше, чем линейными. В случае трансформации клеток E.coli штамм НВ101 плазмидой pBR322 нормальная частота составляет 105-107 трансформантов на 1 мкг ДНК интактной плазмиды.
Очищенную лигированную смесь охладите на льду и добавьте 200 мкл компетентных клеток, инкубируйте препарат 30 мин на льду. Далее прогрейте его в водяном термостате при 420С в течение 3 мин, центрифугируйте в настольной центрифуге 15-20 с и тщательно отберите супернатант. К осадку клеток добавьте 1 мл среды LB и инкубируйте при 370С на качалке 40-60 мин. Клетки осадите центрифугированием и перенесите на селективную среду (твердая среда LB или среда McConkey с конечной концентрацией 100 мкг/мл ампициллина).
Для селекции полученных колоний используют: в случае LB - бело-голубую селекцию (на поверхность среды наносят 25 мкл IPTG (исходная концентрация 20 мг/мл) и 25 мкл X-Gal (20 мг/мл). Если использовать среду McConkey, то клетки наносят прямо на твердую среду.
Электропорация клеток Escherichia coli
Очищенную лигированую смесь охладите на льду и добавьте к 100 мкл компетентных клеток для электропорации, инкубируйте 5 мин на льду, после чего поместите в кювету для электорпорации. Электропорацию проводите при 2,5 В на электропораторе. После пропускания тока к клеткам немедленно добавьте 1 мл нагретой до 37°С LB и поместите на качалку на 37°С на 45-60 мин для начала экспрессии гена устойчивости к ампициллину.
Клетки осадите центрифугированием и перенесите на селективную среду (твердую LB или среду McConkey с конечной концентрацией 100 мкг/мл ампициллина).
Идентификация клонов методом ПЦР с колоний
При клонировании ПЦР-продуктов идентификацию нужных клонов можно проводить с использованием метода ПЦР с колоний. Для этого готовится всё, как в обычном варианте ПЦР, за исключением ДНК. Вместо препарата ДНК вносится фрагмент колонии, который отбирается с поверхности агара с помощью стерильной зубочистки или стерильным наконечником для автоматической пипетки. Режим ПЦР должен соответствовать режиму получения соответствующего ПЦР-продукта.
Еще по теме ПРИЛОЖЕНИЕ 2 Базовые методы генной инженерии:
- Содержание
- ПРИЛОЖЕНИЕ 2 Базовые методы генной инженерии
- Обзор математических методов прогнозирования, особенности использования нечеткой логики принятия решений при мочекаменной болезни