ЦИТОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ПИЩЕВАРЕНИЯ
Аргасовые клещи. Между желудком и отдельными дивертикулами среднего отдела кишечника аргазид в процессе пищеварения обнаруживаются некоторые функциональные различия. Они связаны с наличием у средней кишки двух различных функций.
Последняя одновременно служит и органом пищеварения, и резервным депо больших количеств непереваренной крови. У напитавшихся клещей до удаления коксальной жидкости все отделы средней кишки растянуты до предела. После ее удаления желудок, срединные отростки и, в меньшей степени, основания переднебоковых растянуты кровью значительно слабее, чем дистальные концы переднебоковых и особенно заднебоковые дивертикулы (рис. 270). Последние служат основным местом хранения непереваренной крови и сохраняются в сильно растянутом состоянии даже у голодавших несколько месяцев и способных к повторному питанию клещей (рис. 271, 277). Лишь у очень истощенных особей они запустевают, а их стенки спадаются.Процессы пищеварения первое время после питания сосредоточены преимущественно в желудке и проксимальных частях переднебоковых и срединных отростков. Дистальные концы последних и заднебоковые отростки растянуты в этот период до состояния тонкой мембрановидной оболочки, окружающей массу крови хозяина, и не могут играть существенной роли в ее переваривании. У истощенных особей они, напротив, играют решающую роль в усвоении сохранившихся в них небольших запасов крови.
Форма и размеры эпителиальных клеток средней кишки аргазид сильно варьируют в зависимости от физиологического состояния клеща и, в меньшей степени, от видовой принадлежности. Эпителиальные клетки средней кишки покоятся на тонкой базальной мембране. Снаружи она оплетена сетью взаимно пересекающихся кольцевых и продольных мышечных волокон, не образующих сплошного слоя. Сокращения мускулатуры стенок кишечника обусловливают его постоянные перистальтические движения, хорошо заметные на живых клещах, вскрытых в физиологическом растворе.
По морфологическим особенностям, характеру внутриклеточных включений и функциональной активности, мы различаем три типа клеток: секреторные, пищеварительные и недифференцированные резервные, сходные с таковыми скорпионов (Павловский и Зарин, 1926), пауков (Millot, 1949; Иванов, 1965) и изученных в этом отношении групп клещей (Bader, 1938; Hughes, 1950, 1952).
Соотношение между тремя описанными типами клеток до сих пор не вполне ясно.
Несомненно, что пищеварительные и секреторные клетки образуются из резервных. У неполовозрелых фаз их массовое размножение обычно наблюдается вскоре после питания при подготовке к линьке. У половозрелых особей митотические деления недифференцированных клеток встречаются значительно реже, но обычно также связаны с окончанием кровососания. Скорее всего у аргасовых клещей существует один тип резервных клеток, из которых в зависимости от фазы пищеварения дифференцируются в большем числе секреторные или пищеварительные клетки. В то же время возможность прямого взаимного превращения двух последних типов после завершения их дифференцировки, как считает Р. Татчелл (Tatchell, 1964), маловероятна. Еще менее правдоподобно предположение того же автора о возможности развития клетки противоположного типа из сохранившегося после отторжения апикальной части проксимального конца клетки. По нашим наблюдениям, апикальные концы клеток чаще всего отрываются в просвет вместе с участком цитоплазмы, включающим ядро, тогда как сохранившаяся проксимальная часть клетки вскоре дегенерирует.Наиболее детально нами были исследованы цитологические особенности кишечника самок Ornithodoros papillipes. У голодных клещей, готовых к питанию, но еще сохранивших в полости средней кишки значительное количество полупереваренной крови от прошлого кровососания, в стенках кишечника преобладают клетки секреторного и резервного типов. Секреторные клетки выдаются апикальными концами в просвет и имеют столбчатую форму при высоте 60—120 мк и максимальной ширине 20—30 мк (рис. 281). Овальные или почти правильные сферические ядра диаметром 6—7 мк лежат посередине или несколько ближе к проксимальному концу клетки. Проксимальнее ядер цитоплазма мелкозернистая, с тонкими фибриллярными структурами, по-видимому, представляющими элементы эндоплазматической сети, и большим количеством митохондрий. В апикальной половине клетки располагаются вакуоли, обычно не красящиеся ни кислыми, ни основными красителями. Их содержимое дает интенсивную ШИК-положительную реакцию, сохраняющуюся после обработки срезов диастазой.
По характеру гистохимических реакций их содержимое, вероятно, является полисахаридом, тогда как более мелкие ШИК-положительные гранулы в проксимальной цитоплазматической зоне — гликогеном.Резервные клетки более мелкие, и их форма приближается к кубической или клиновидной. Их дистальные концы часто прикрыты смыкающимися сверху соседними пищеварительными или секреторными клетками (рис. 280, 281). Пищеварительные клетки у голодающих особей относи-
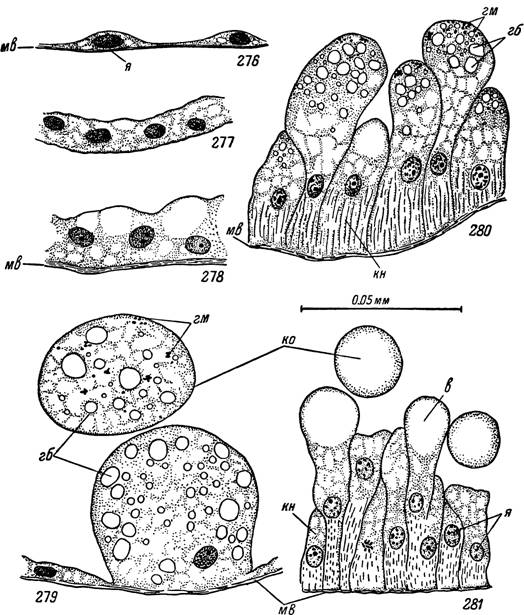
Рис. 276—281. Строение клеток средней кишки самки Ornithodoros
papillipes.
276 — участок заднебокового отростка сразу после питания; 277 — тоже через несколько месяцев голодания; 278 — стенка желудка в растянутом состоянии; 279 — отшнуровывание глыбовидных пищеварительных клеток в полость кишечника; 280 — пищеварительные клетки и 281 — секреторные клетки через несколько месяцев после кровососания.
я — ядра; мв — мышечные волокна; ко — клетки, оторвавшиеся в просвет; в — вакуоли с секретом; гб — включения гемоглобина; гм — включения гематина; кн — недифференцированные эпителиальные клетки.
тельно немногочисленны и разбросаны вперебежку с секреторными и резервными. Они имеют вытянутую форму с расширенной апикальной половиной или же почти кубическую, но значительно крупнее резервных и с выдающейся в просвет апикальной поверхностью. Ядра занимают центральное положение, а в цитоплазме содержится большое количество различных включений (рис. 279, 280). Среди последних в апикальной половине клетки преобладают вакуоли диаметром от 1 до 3—6 мк, крася-
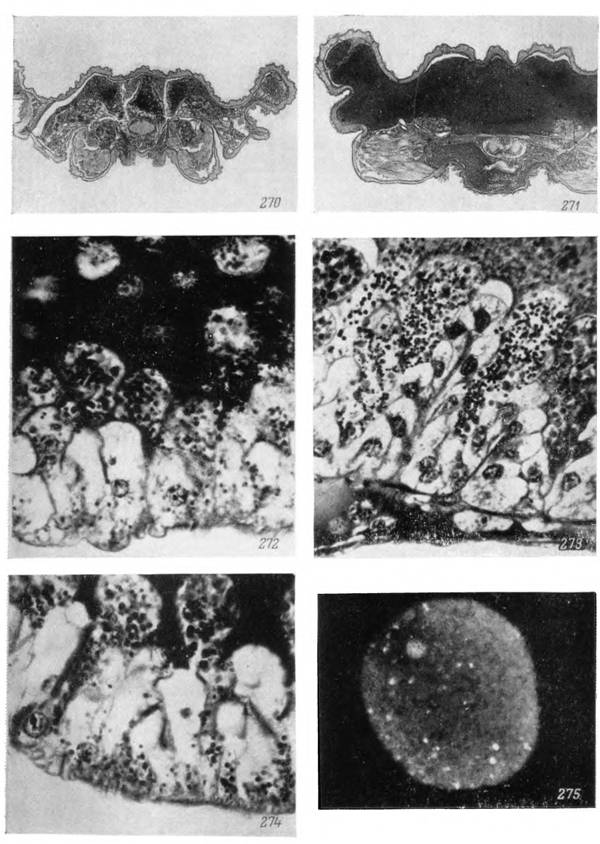
Рис. 270—275. Строение средней кишки аргасовых клещей.
Поперечные срезы: 270 — через голодную самку Ornithodoros papillipes, 271 — то же через напитавшуюся, 25; участки стенки средней кишки самок: 272 — О. papillipes, 273 — Alveonasus lahorensis, 274 — Argas persicus, X600; 275 — отторгнутая пищеварительная клетка Ornithodoros papillipes, прижизненная подкраска акридиновым оранжевым, Х900, люминесцентный микроскоп.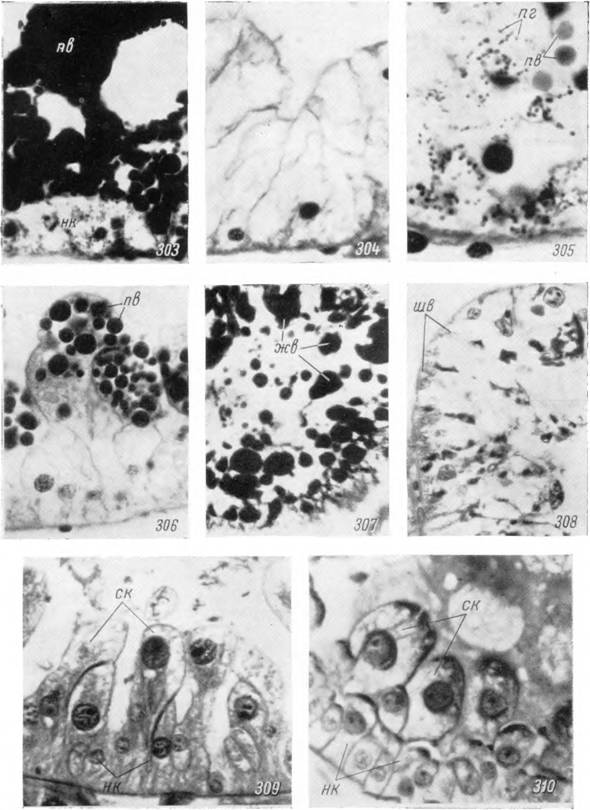
Рис.
303—310. Клетки средней кишки самки Hyalomma asiaticum.303 — голодная сразу после линьки; 304 — истощенная, через 8 месяцев после линьки, окраска железным гематоксилином-эозином; 305 — гранулы пурпурного пигмента в клетках голодной особи; 306 — пищевые включения из гемоглобина (окраска бен- зидином-сафранином); 307 — жировые вакуоли (окраска черным Суданом);
308 — ШИК-положительные вещества в клетках кишечника голодного клеща;
309 — секреторные клетки кишечника во время дифференцировки; 310 — то же в период массового отторжения в полость кишечника в начале питания (окраска ШИК- гематоксилин Эрлиха). Микрофотографии: 305 — X 1350; 307 — Х400; 303, 304, 306,
308—310 — Х600.
пв — пищевые включения из гемоглобина; же — жировые вакуоли; шв — ШИК-положителыюе вещество; пг — пигментные гранулы; нк — недифференцированные клетки; ск — секреторные клетки.щиеся азокармином и железным гематоксилином. Они дают интенсивную положительную реакцию на белок с БФС и на гемоглобин с бензидином, так что их следует считать гемоглобином крови хозяина, поглощенным в мало измененной форме клеткой клеща.
Второй тип включений представлен мелкими (до 1 мк) и средними гранулами гематина, разбросанными по всей цитоплазме. Более крупные гематиновые включения (2—4 мк) имеют форму правильных сферокристал- лов интенсивной буро-черной окраски. При обработке срезов методом ШИК или кармином по Бесту в цитоплазме выявляется также очень большое количество мелких (до 1 мк) гранул гликогена, а на замороженных
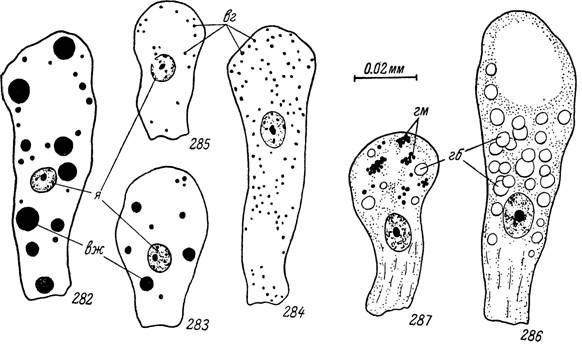
Рис. 282—287. Типы включений в пищеварительных клетках Ornithodoros papillipes. 282, 284, 286 — недавно напитавшиеся особи; 283, 285, 287 — истощенные особи.
я — ядра; вж — вакуоли с широм; вг — включения гликогена; гб — гемоглобин;
гм — гематин.
срезах после окраски Суданом черным или III — средних и крупных жировых вакуолей (рис. 282—287). Последние также интенсивно чернятся при осмировании. Среди пищеварительных клеток, сильно загруженных гематином, часто наблюдается их отторжение в просвет кишечника, где они округляются и приобретают сходство с амебоцитами (рис.
275, 279). В дальнейшем они подвергаются полной дегенерации, а гематин смешивается с содержимым кишечника.При заполнении кишечника свежей кровью наблюдается массовое разрушение секреторных клеток, а содержащееся в них полисахаридное вещество освобождается в просвет; часть секреторных клеток при этом сперва отшнуровывается и лишь затем подвергается растворению (рис. 281). Резервные и пищеварительные клетки могут очень сильно растягиваться и превращаются в тонкую мембрану с небольшими утолщениями в местах нахождения ядер, границы между отдельными клетками становятся неразличимыми (рис. 276, 278). Истончение стенки кишечника местами достигает такой степени, что создаемся впечатление, что она образована одной базальной мембраной, опирающейся на мышечное сплетение.
Постепенно растяжение желудка и проксимальных концов передне- боковых отростков уменьшается, а клетки начинают выдаваться в просвет. В первое время после питания основную массу клеток составляют пищеварительные. Вначале они имеют глыбовидную или кубическую форму.
Группы подобных клеток высотой в 30—50 мк обычно бывают окружены уплощенными участками (рис. 279). Очень часто эти клетки целиком отрываются в просвет, формируя крупные сферы с ядрами. Число кубических клеток продолжает увеличиваться, они вытягиваются внутрь полости кишечника и форма их приближается к столбчатой. Параллельно все в большем количестве появляются секреторные клетки с вакуолизи- рованными дистальными концами.
Внутри пищеварительных клеток начинают появляться мелкие вакуоли (до 1 мк), дающие положительную реакцию с БФС и бензидином. При их слиянии образуются более крупные пищевые вакуоли, уже описанные у голодающих особей. Вакуоли с гемоглобином могут заполнять значительную часть апикальной цитоплазматической зоны. В дальнейшем они начинают исчезать, а на их месте формируются буро-черные конкреции гема тина. Внутриклеточное переваривание гемоглобина не идет синхронно во всех вакуолях одной клетки, так что в цитоплазме одновременно встречаются как крупные пищевые вакуоли, так и конечный продукт переваривания их содержимого — гематин.
Важно также отметить, что с началом внутриклеточного переваривания гемоглобина в клетке перестают появляться новые пищевые вакуоли, а по окончании переваривания всех вакуолей пищеварительные клетки сильно загружаются гематином и большей частью подвергаются дегенерации.Поглощение огромных молекул гемоглобина происходит главным образом путем пиноцитоза, но недостаточная разрешающая способность светового микроскопа не позволяет провести непосредственные наблюдения этого явления. В результате о существовании пиноцитоза в пищеварительных клетках мы может судить лишь по его конечным результатам, т. е. по появлению в цитоплазме вакуолей с гемоглобином. Прохождение их сквозь цитоплазматическую мембрану невозможно объяснить на основании нормального механизма клеточной проницаемости. Определенную роль в поглощении гемоглобина, а иногда и целых эритроцитов играют процессы истинного фагоцитоза. На поверхности пищеварительных клеток при прижизненных наблюдениях иногда удается наблюдать образование нитевидных или даже лопастевидных отростков, вытянутых в просвет. Сближаясь своими дистальными концами, они охватывают часть содержимого кишечника, так что оно оказывается заключенным во внутриклеточную вакуоль.
На апикальной цитоплазматической мембране иногда выявляется узкая наружная зона с поперечной исчерченностыо, ширина которой обычно не превышает 1 мк. Татчелл (Tatchell, 1964) наблюдал аналогичное образование в клетках Argas persicus и считает его зоной микроворсинок. Окончательные доказательства существования микроворсинок в пищеварительных клетках аргазид, однако, могут быть получены только с помощью электронной микроскопии.
Наряду с гемоглобином пищеварительные клетки усваивают и липоидное вещество стром эритроцитов. В цитоплазме клеток на этой стадии
Рис. 288—298. Участки кишечника в разные периоды переваривания крови личинками и нимфами Alveonasus lahorensis и Ornithodoros papillipes.
Alveonasus lahorensis: 288 — голодные личинки; 289 — 2 суток прикрепления; 290 — полностью напитавшиеся; 291 — через 3 суток по окончании кровососания; 292 — нимфы III возраста вскоре после линьки; 293 — к концу питания; 294 — через 3 суток после окончания питания. Ornithodoros papillipes: 295 — голодные личинки; 296 — напитавшиеся личинки; 297 — голодные нимфы I возраста; 298 — напитавшиеся нимфы.
пк — пищеварительные клетки; нк — недифференцированные клетки; мт — митотические деления клеток стенки кишечника; on — отторгнутые пищеварительные клетки в полости кишечника; э — эритроциты крови хозяина; гб — внутриклеточные включения гемоглобина; гм — гематин; пгб — кристаллы гемоглобина в полости кишечника.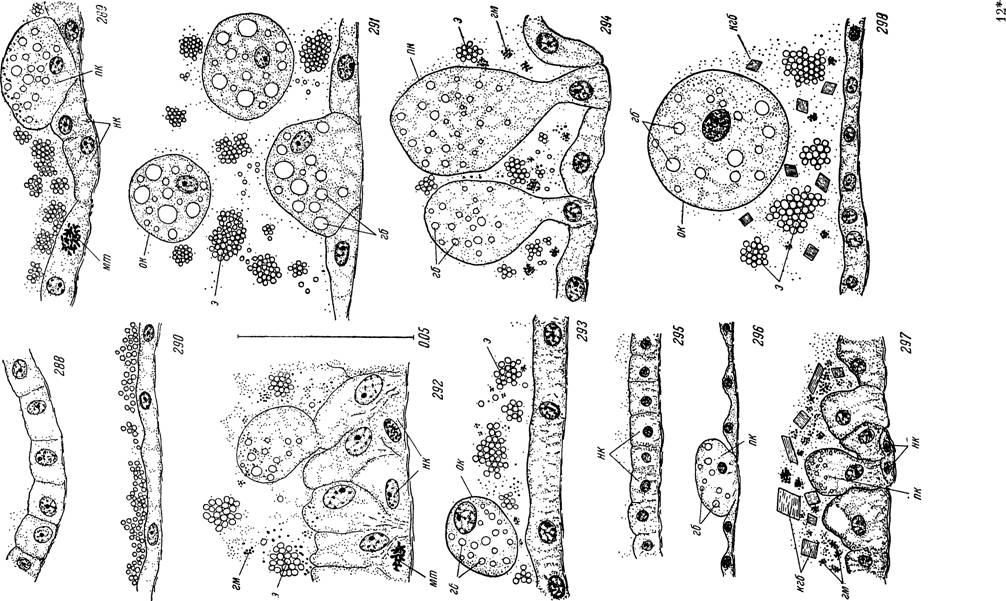 накапливаются липоидные вакуоли (рис. 282). Последнее, однако, не исключает и возможность их формирования путем дезаминирования аминокислот белков крови хозяина в цикле клеточного метаболизма.
накапливаются липоидные вакуоли (рис. 282). Последнее, однако, не исключает и возможность их формирования путем дезаминирования аминокислот белков крови хозяина в цикле клеточного метаболизма.
Весь описанный цикл изменений свойствен не только клеткам, прикрепленным к базальной мембране, но и их ядерным амебовидным фрагментам, плавающим в содержимом кишечника. На основании этого можно предположить, что отторжение клеток средней кишки связано не только с завершением их цикла развития и началом дегенерации, но и имеет прямое отношение к механизму переваривания крови.
Описанная схема справедлива для нимф и имаго Ornithodoros papil- lipes. Несколько отличаются лишь гистологические изменения в кишечнике личинок. У голодных личинок он построен из небольших, почти кубических клеток, слабо выдающихся в просвет. В пищеварительных клетках в этот период неразличимы какие-либо включения и внешне они не отличаются от секреторных (рис. 295). У напитавшихся клещей стенки кишечника бывают очень сильно растянуты (рис. 296). Размеры отдельных клеток постепенно увеличиваются, они вдаются в просвет, а часть их открывается и свободно плавает в содержимом кишечника. Внутри клеток появляются многочисленные пищевые включения. Параллельно идет очень быстрое размножение недифференцированных клеток, превращающихся в дальнейшем в секреторные и пищеварительные. Деление клеток всегда только митотическое. У перелинявших нимф I гистологическое строение кишечника уже такое же, как и у взрослых особей (рис. 297).
У быстро насасывающихся нимф и имаго Argas persicus, Ornithodoros moubata, а также взрослых Alveonasus lahorensis гистологические изменения кишечника при пищеварении в основных чертах сходны с Ornithodoros papillipes, хотя имеются и определенные отличия.
Для Argas persicus характерна значительно большая вытянутость эпителиальных клеток, особенно у голодных клещей. Проксимальные концы клеток собраны в многочисленные мелкие складки и отличаются значительной фибриллярной исчерченностью (рис. 274). У истощенных клещей кишечник, как правило, содержит один гематин, его стенки спадаются и просвет исчезает. Интересная особенность пищеварения A. persicus связана с питанием птичьей кровью. Ядерные эритроциты птиц весьма устойчивы к гемолизу и многие из них непосредственно фагоцитируются пищеварительными клетками, так что здесь мы имеем классический пример внутриклеточного переваривания целой, неповрежденной клетки.
У Ornithodoros moubata в связи с отсутствием сообщения средней кишки с ректальным мешком и анальным отверстием дистальные концы клеток и полость кишечника обычно очень сильно загружены гематином, лишенным выхода наружу (рис. 272).
У голодных половозрелых Alveonasus lahorensis стенки кишечника собраны в правильную систему. складок, придающих ему характерную морщинистую внешность (рис. 273). Во время кровососания за счет их расправления объем кишечника значительно увеличивается. Параллельно с этим происходит и сильное растяжение клеток. Процессы пищеварения происходят значительно медленнее, чем у Ornithodoros papillipes и особенно у Argas persicus.
У питающихся несколько суток личинок и нимф Alveonasus lahorensis гистологические изменения кишечного эпителия при пищеварении очень сходны с одно- и двуххозяинными иксодовыми клещами. У голодных личинок кишечник построен из однородных кубических клеток, бедных включениями (рис. 288). С началом кровососания они вытягиваются в просвет, а в их дистальных концах появляются секреторные вакуоли и* пищевые включения (рис. 289). Часть клеток или их дистальные концы отшнуровываются и плавают в просвете. Примерно со вторых суток питания среди резервных клеток наблюдается значительное количество митотических делений. К концу питания заметно выросший за счет размножения клеток кишечник начинает растягиваться (рис. 290). В его полость вдаются лишь отдельные глыбовидные пищеварительные клетки (рис. 291). Число их увеличивается, они смыкаются и принимают нормальную вытянутую форму. За короткое время подготовки к линьке на нимф I, а затем на нимф II и III успевает перевариваться лишь незначительная часть крови, поглощенной на предшествующих стадиях. Однако клещи сразу же после окончания линьки вновь приступают к кро- вососанию, так что свежая кровь смешивается с непереваренной старой и конечными продуктами пищеварения. Стенки кишечника нимф в зависимости от степени растяжения выдаются в просвет (рис. 294) или уплощены (рис. 293). Почти всегда удается обнаружить значительное количество митозов, которые происходят в недифференцированных клетках, прилегающих к базальной мембране. Таким образом, с момента прикрепления личинок и до отпадения напитавшихся нимф III происходит непрерывное переваривание крови хозяина. Многодневные периоды кровососания чередуются с короткими периодами линьки. Во время кровососания кровь поглощается относительно небольшими порциями и часть ее сразу же переваривается. Поэтому растяжение стенок всегда слабее, чем у быстра питающихся видов.
Иксодовые клещи. У иксодовых клещей с однократным питанием на каждой фазе развития циклические изменения в строении кишечного эпителия выражены резче, чем у аргазид. Они связаны не только с переходом от состояния относительного «покоя» при голодании к интенсивной функциональной активности во время питания и при подготовке к линьке или яйцекладке, но также определяются физиологическим возрастом голодных клещей или периодом питания.
В связи со сложностью и разнообразием гистологических изменений во время пищеварения описание этих процессов приводится отдельно для самок, самцов, личинок и нимф Нуаіотта asiaticum и Ixodes ricinus, а в необходимых случаях рассматриваются наиболее существенные отклонения у других видов.
У иксодовых клещей процессы пищеварения протекают интенсивнее, чем у аргазид, так как поглощенная пища в первые дни питания полностью захватывается пищеварительными клетками, а их гипертрофированные дистальные концы заполняют значительную часть просвета кишечника (рис. 300). В результате различия в характере пищеварения в центральной части кишечника и боковых дивертикулах практически не выражены или же связаны с различной степенью заполнения пищей и растяжением стенок отдельных отростков с начала питания до стадии растяжения. В период растяжения быстрое увеличение размеров клеща происходит в основном за счет растягивания йокровов тела и стенок средней кишки. Полость кишечника заполняется массой непереваренной крови хозяина, а клетки стенок необычайно уплощаются или же отторгаются в просвет. Подобная крайняя степень растяжения свойственна всем частям средней кишки полностью напитавшейся особи (рис. 301). В дальнейшем на стадии линьки или яйцекладки растяжение стенок кишечника заметно ослабевает за счет усвоения части поглощенной пищи, а просвет почти полностью занят гипертрофированными пищеварительными клетками (рис. 302).
Самки. Гистологические различия между отдельными дивертикулами практически отсутствуют, но в зависимости от степени заполнения клеток запасами питательных веществ у голодных особей или полости кишечника пищей во время питания наблюдаются заметные различия в их строении, связанные с нахождением на разных стадиях пищеварения.
У недавно перелинявших самок Hyalomma asiaticum строение средней кишки мало отличается от нимф в конце линьки. Гипертрофированные пищеварительные клетки, сохранившиеся от нимфальной фазы, достигают 200— 260 мк высоты и обычно занимают весь просвет (рис. 303 на стр. 177). Их цитоплазма заполнена сферическими пищевыми включениями диаметром от 2 до 20 мк. Они полупрозрачны и имеют естественную желтовато-коричневую окраску. Железный гематоксилин окрашивает их в черный цвет, а эозин — в розовый. При тройной окраске азокармином они в большинстве случаев красные, но часть из них всегда синяя. Между ними лежат мелкие сферические включения гематина диаметром в 1—2 мк буро-черного цвета. Границы между отдельными клетками на этой стадии обычно неразличимы, а ядра часто маскируются описанными включениями. В гипертрофированных клетках они неправильной многоугольной формы диаметром 10—12 мк. Ядра недифференцированных клеток, лежащие вблизи базальной мембраны, круглые или овальные и меньшего размера (5 мк). К концу стадии послелиночного доразвития количество пищевых включений заметно уменьшается, гипертрофированные клетки спадаются, а часть их разрушается и стенки кишечника приобретают нормальный для голодных клещей вид.
У особей, недавно закончивших послелиночное доразвитие, среди столбчатых эпителиальных клеток высотой в 30—50 мк встречаются отдельные гипертрофированные клетки, целиком заполненные пищевыми включениями. Однако большая часть пищеварительных клеток, сохранившихся от нимфальной фазы, содержит пищевые включения диаметром 2—20 мк преимущественно в апикальных частях, а ядра размером в 5— 6 мк занимают базальные части цитоплазмы (рис. 306). Наиболее многочисленны у голодных особей недифференцированные клетки. Кроме того, встречаются и типичные секреторные клетки. Их апикальные части выдаются в полость кишечника, сильно вакуолизированы и заполнены ШИК-положительным веществом. Внутри отростков всегда существует достаточно обширная полость. Она бывает пустой или же содержит кристаллики гематина и фрагменты разрушенных клеток.
В процессе голодания гипертрофированные пищеварительные клетки полностью исчезают и белковые пищевые включения выявляются в немногих сохранившихся пищеварительных клетках (рис. 304). В то же время в их цитоплазме очень многочисленны включения гематина. Цитоплазма подобных клеток выглядит оптически пустой и очень слабо окрашивается эозином или световым зеленым. В стенках средней кишки образуются многочисленные складки, а в ее полости увеличивается масса из гематина и остатков клеток. Внешний вид недифференцированных клеток остается прежним.
У истощенных особей отростки кишечника обычно спадаются. Внутри их сильно суженного просвета местами встречаются очень крупные скопления гематина. Пищеварительные клетки становятся крайне редкими. В их цитоплазме отсутствуют пищевые включения, но много гематина.
У самок Ixodes ricinus строение кишечника у голодных особей в основных чертах сходно с описанным для Hyalomma asiaticum. У только что перелинявшего клеща клетки кишечника заполнены вакуолями с пищевыми включениями, размеры которых сильно варьируют. Вакуоли заключены в нормальных и в гипертрофированных пищеварительных клетках, сохранившихся от нимфальной фазы, но границы между отдельными клетками, как и их ядра, в первые дни после линьки обычно не видны. Лишь на более поздних стадиях, по мере потребления пищевых включений,
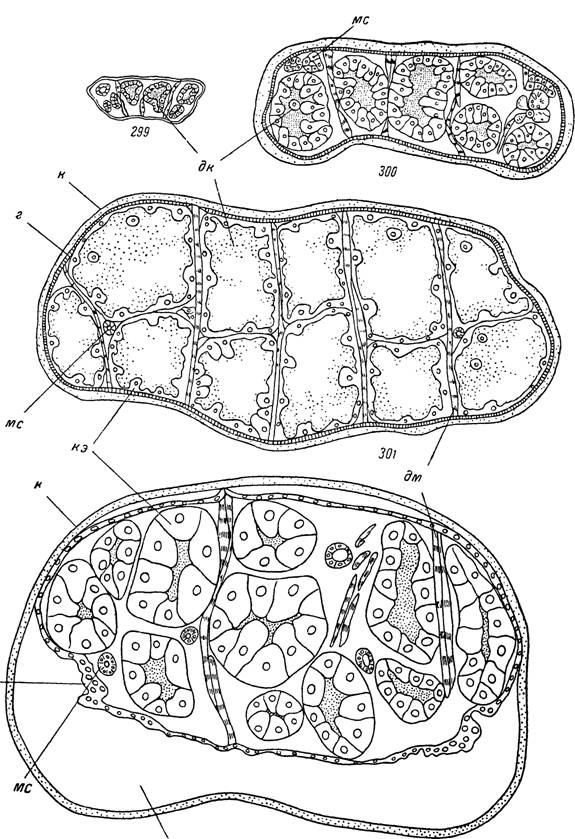
299 - голодная-
e _ . Сразу по отпаДении; 302 - в середине стадиГлиньки.ЫаПИТаВШаЯСЯ
клетки стенки средней^мшки; —^альтогиев™сосудьгЄаж ™!?ooc' ** ~ эпителиальные
лп - ливдадм полость ’ ~ Д°Рсо-вентральные мышцы;
начинают выступать границы отдельных гипертрофированных клеток. Они обычно находятся на различных стадиях дегенерации. Между пищевыми включениями внутри клеток и в полости кишечника заключено большое количество буровато-черных мелких гранул гематина. Последний придает среднему отделу кишечника клещей характерную бурую или почти черную окраску.
К концу стадии послелиночного доразвития пищевые резервы в клетках резко сокращаются, а значительная часть гипертрофированных клеток, сохранившихся от нимфальной фазы, подвергается разрушению. У голодных, способных к нападению клещей стенки средней кишки построены из столбчатых или почти кубических недифференцированных клеток высотой 10—20 мк с небольшими сферическими ядрами и бедной включениями цитоплазмой (рис. 318, 119). Местами между ними вклиниваются более крупные (высота 30—40 мк) булавовидные секреторные клетки с сильно вакуолизированной цитоплазмой и пищеварительные клетки нимфальной фазы. Количество последних зависит от времени, прошедшего после линьки. В цитоплазме нимфальных клеток содержатся сравнительно крупные пищевые включения и значительное количество мелких гранул гематина. В отличие от Н. asiaticum недифференцированные клетки Ixodes ricinus имеют более правильную столбчатую форму и резко выраженные межклеточные границы. В сохранившихся пищеварительных клетках всегда скапливается большое количество гематина. Различия между недифференцированными и секреторными клетками выражены резче.
Изучение динамики резервных белков, гликогена и жиров у обоих видов показывает, что описанные нами гистологические изменения объективно отражают процессы расходования запасных питательных веществ, накопленных на нимфальной фазе. Красящиеся железным гематоксилином, эозином и азокармином сферические включения диаметром от 2 до 20 мк приобретают интенсивную сине-фиолетовую окраску при обработке бромфеноловым синим в сулеме. Это указывает на их белковую природу. Вместе с тем они всегда дают положительный результат при бензидиновой реакции на гемоглобин (рис. 306). В пользу того, что они представляют мало измененный гемоглобин крови хозяина, говорят и косвенные данные. При их внутриклеточном переваривании в цитоплазме увеличивается содержание гематина — конечного продукта расщепления гемоглобина членистоногими. Интенсивность окраски различными гистологическими красителями или бромфеноловым синим неодинакова во всех описываемых включениях. Последнее, вероятно, отражает различные этапы их переваривания и усвоения. Следует отметить, что подобные включения, кроме гемоглобина, вероятно, содержат и другие вещества. В их состав входят мукополисахариды. Они дают слабую положительную реакцию на углеводы по методу ШИК и пурпурно-фиолетовую метахрома- зию при обработке толуидиновым синим.
В клетках средней кишки находятся значительные запасы гликогена. После обработки по методу ШИК внутри клеток кишечника выявляется значительное количество мелких (до 1 мк диаметром) красных гранул, отсутствующих на контрольных срезах, обработанных амилазой. Очень часто в дистальных концах клеток они образуют крупные скопления. Последние, вероятно, не отражают истинную локализацию гликогена и образуются вследствие диффузионных токов при фиксации (рис. 308).
При обычных методах фиксации и заливки в парафин вследствие обезвоживания и экстракции липоидов клетки кишечника сильно сжимаются, а внутри них появляются многочисленные оптически пустые пространства. Фиксация осмием (смесь Шампи) не устраняет отмеченные недостатки, хотя в клетках и сохраняется часть чернящихся жировых вакуолей.
Только при фиксации формалином и изготовлении срезов на замораживающем микротоме сжатие сводится до минимума и экстракция липоидов отсутствует. Пищеварительные клетки при этой методике выглядят упругими, а на месте прозрачных участков мы видим многочисленные жировые вакуоли (рис. 307). Подобные вакуоли интенсивно красятся Суданом черным или III и занимают значительную часть цитоплазмы. Размеры их колеблются от 4 до 40 мк (основная масса 10—20 мк).
Содержание рибонуклеиновой кислоты в недифференцированных клетках голодных клещей относительно невелико и не проявляет заметных колебаний. Последнее, вероятно, связано с отсутствием процессов роста и органогенеза в этот период. В пищеварительных клетках содержание плазматической РНК ничтожно и она выявляется с большим трудом.
Вариации в длительности периода послелиночного доразвития и способности к голоданию связаны с различным количеством запасных питательных веществ и скоростью их расходования у разных видов. Самки Ixodes ricinus и Hyalomma asiaticum обладают продолжительным периодом доразвития, большой устойчивостью к голоданию и соответственно очень большим количеством запасных питательных веществ. У быстро развивающихся Н. anatolicum, Boophilus calcaratus и Rhipicephalus turanicus пищевых включений значительно меньше. Совсем мало их у довольно эфемерных личинок и нимф рода Dermacentor. У нападающих самок в кишечнике содержится еще значительное количество пищевых веществ, расходуемых по мере дальнейшего голодания. Лишь после очень длительного голодания у самок Ixodes ricinus, в наших опытах достигавших 1.5 лет, кишечник совершенно пустой. У Dermacentor pictus, способного, поданным Н. Г. Олсуфьева (1953), к активности в продолжение двух и более сезонов, мы обнаружили особенно значительное количество клеток с пищевой зернистостью в кишечнике нападающих самок.
В отношении клеточного состава средней кишки иксодид мнения до сих пор расходятся. Ряд исследователей считает, что уже у голодных особей в ее состав входят три типа клеток: недифференцированные резервные, дегенерирующие пищеварительные и секреторные (Roesler, 1934; Hughes, 1954). С другой стороны, распространено мнение, что стенка кишечника построена из одного типа клеток, форма и строение которых могут очень варьировать в зависимости от стадии пищеварения (Till, 1961; Chinery, 1964). На основании изучения цитологических и гистохимических особенностей эпителия средней кишки мы считаем, что все входящие в его состав клетки имеют общее происхождение. Исходным типом, как и у аргасовых клещей, мы считаем недифференцированные резервные клетки, причем у голодных особей этот тип клеток наиболее многочислен, а секреторные клетки встречаются реже. Пищеварительные клетки, заполненные различного типа включениями, образуются в результате дифференцировки резервных клеток. При этом специализация в направлении образования секретов для полостного пищеварения («секреторные клетки») или накопления в цитоплазме и внутриклеточного переваривания крупных белковых или жировых вакуолей («пищеварительные клетки), вероятно, имеет необратимый характер и, закончив цикл функциональной активности, клетка подвергается дегенерации. Тем не менее исключения не столь редки, так как нам неоднократно приходилось наблюдать появление в клетках с крупными секреторными вакуолями включений с гемоглобином, т. е. превращение секреторных клеток в пищеварительные. Таким образом, с определенными оговорками мы также признаем существование в среднем отделе иксодовых клещей трех типов клеток: недифференцированных резервных, пищеварительных и секреторных, гомологичных таковым аргасовых клещей.
Для иксодид характерна определенная физиологическая недоразвитость эпителия средней кишки у голодных особей. В нем еще отсутствуют готовые к функционированию пищеварительные клетки, тогда как пищеварительные клетки от нимфальной фазы служат депо запасных питательных веществ и не способны к участию в новом пищеварительном цикле.
Сразу же после прикрепления к хозяину происходит быстрый рост секреторных клеток. Отдельные недифференцированные клетки также начинают увеличиваться в размерах, их дистальные концы выдаются в просвет и они превращаются в клетки секреторного типа (рис. 309 и 320). В отличие от еще сохраняющихся местами пищеварительных клеток нимфальной фазы, в сильно вакуолизированной цитоплазме секреторных клеток отсутствуют пищевые включения и гранулы гематина. В то же время в их апикальной цитоплазматической зоне появляется большое количество ШИК-положительного вещества в форме мелких гранул (менее 1 мк), местами образующих довольно плотные скопления в пространстве между прозрачными вакуолями. Эта зернистость сохраняется и на срезах, обработанных диастазой, так что ее следует считать полисахаридом, отличным от гликогена.
Повышение содержания РНК в резервных клетках — первый признак начала их дифференцировки. Последняя обнаруживается еще до поступления жидкой пищи в полость кишечника («подготовительный период питания»). На срезах через клещей в первые часы после прикрепления местами хорошо видны отдельные сильно увеличенные вакуолизирован- ные клетки, вдающиеся в пустой, не содержащий никаких следов пищи кишечник. В дальнейшем образование новых секреторных клеток происходит на протяжении всего времени питания. В первые 1—2 дня после прикрепления количество секреторных клеток еще сравнительно невелико и они не образуют сплошного слоя, а как бы вклиниваются в толщу недифференцированных резервных. По мере усвоения пищи их число резко увеличивается, они тесно соприкасаются дистальными концами и как бы создают второй клеточный слой над лежащими базальнеє резервными клетками. Размеры последних существенно не меняются по сравнению с таковыми у голодных особей (высота 10—15 мк).
Секреторные клетки могут достигать весьма значительных размеров: у Hyalomma asiaticum их высота составляет 70—100 мк при ширине в 25— 30 мк в дистальной части, а у Ixodes ricinus — 50—70x30—40 мк. Форма клеток чаще всего бывает булавовидной или столбообразной, но с сужением к базальному концу. На первых этапах дифференцировки они чаще бывают глыбовидными. Ядра располагаются ближе к середине клетки. В то время как в апикальной половине цитоплазма сильно вакуолизиро- вана, в базальной половине она содержит большое количество базофиль- ных волокнистых структур. При окраске метиловым зеленым—пирони- ном последние дают интенсивную положительную реакцию на присутствие РНК и, по-видимому, их следует считать элементами эргастоплазмы. Параллельно с увеличением содержания РНК происходит значительный рост ядер, достигающих у Hyalomma asiaticum диаметра 15—20 мк и у Ixodes ricinus — 13—20 мк против 7—9 мк в недифференцированных клетках. Форма ядра при этом может становиться неправильно многоугольной. Рибонуклеиновое ядрышко также значительно увеличивается и часто фрагментируется на 2—3 части. Параллельно с ростом ядра уменьшается интенсивность его окрашивания по Фельгену, а на конечных этапах развития они начинают окрашиваться кислыми красителями и перестают давать специфические реакции на ДНК.
С поступлением первых порций жидкой пищи в полость кишечника начинается разрушение дистальных концов или отторжение целых клеток в просвет кишки (рис. 310 и 321). Ядерные фрагменты при этом округляются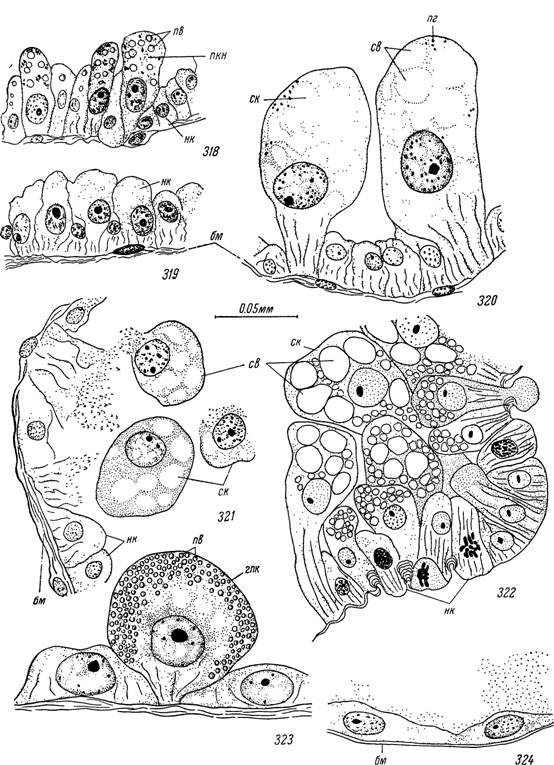
Рис. 318—324. Недифференцированные, секреторные и пищеварительные клетки самки Ixodes ricinus на стадии активности и во время питания.
318 — голодная особь в начале и 319 — в конце стадии активности; 320 — дифференцировка секреторных клеток; 321 — отторжение секреторных клеток в полость кишечника в начале питания; 322 — «крипта» с делящимися недифференцированными клетками; 323 — гипертрофированная пищеварительная клетка; 324 — растянутый участок стенки бокового отростка.
нк — недифференцированные клетки; пкн — пищеварительные клетки нимфальной фазы; ск — секреторные клетки; св — вакуоли с секретом; бм — базальная мембрана и мышечные волокна; пв — пищевые включения; пг — пигментные гранулы; гпк — гипертрофированные пищеварительные клетки.и приоб ретают некоторое сходство с гемоцитами. В дальнейшем они, однако, также подвергаются разрушению. Содержимое секреторных .клеток при этом смешивается с пищей, так что в белковой массе последней обнаруживаются как бы островки ШИК-положительного коллоидного вещества. Ряд исследователей (Hughes, 1954; Chinery, 1964) считает, что полисахаридный коллоидный секрет обеспечивает гемолиз эритроцитов. Процесс отторжения секреторных клеток может идти столь интенсивно, что местами на базальной мембране сохраняются лишь отдельные изолированные группы резервных клеток и может создаться впечатление, что пища отграничена от полости тела только тонкой базальной мембраной.
В первые дни после прикрепления поглощаются относительно небольшие количества пищи, большая часть ее сразу же усваивается клещом, так что заметного растяжения стенок кишечника обычно не наблюдается. В то же время диаметр дивертикулов сильно увеличивается и это происходит несмотря на непрерывное разрушение значительного числа секреторных клеток. Источником замещения разрушающихся клеток и роста дивертикулов служит интенсивное размножение недифференцированных клеток. Митотические деления наблюдаются уже на вторые сутки прикрепления и в продолжении всего времени питания, но особенно многочисленны они в начале периода роста клеща (рис. 313 на стр. 192). Сперва размножение клеток происходит по всей поверхности кишечника, но примерно с середины периода роста оно особенно часто наблюдается в складках между гипертрофированными специализированными клетками (рис. 322). Эти складки внешне очень сходны с криптами насекомых и представляют собой скопления недифференцированных эпителиальных клеток, глубоко вдающихся в полость тела сквозь редкое сплетение мускульных волокон.
Переваривание и усвоение жидкой пищи начинается сразу же по поступлении первых ее порций в полость кишечника. Однако в первые дни питания эти процессы не сопровождаются появлением крупных пищевых включений или продуктов их разрушения в цитоплазме клеток. Тем не менее процессы пищеварения уже с самого начала питания протекают с большой интенсивностью. О переваривании и адсорбции белкового содержимого средней кишки в этот период можно судить по изменению его окраски в узкой зоне, граничащей с поверхностью клеток, и по появлению мелких белковых гранул (диаметром менее 1 мк) в периферической зоне цитоплазмы сравнительно крупных глыбовидных клеток, встречающихся в стенке кишки наряду с секреторными и резервными. К сожалению, доступные гистохимические методики не позволяли выявить более тонкие изменения, связанные с разрушением и всасыванием белков, отличных от гемоглобина. Косвенным доказательством эффективного использования поступающей пищи было в основном очень быстрое увеличение размеров тела питающегося клеща, связанное с ростом кутикулы идиосомы, гонады, кишечника и других внутренних органов.
Гистологические изменения, связанные с поступлением в клетки и внутриклеточным перевариванием белков, четко прослеживаются лишь при питании кровью. Гемоглобин из разрушенных эритроцитов хозяина легко выявляется бензидиновой реакцией. При его переваривании образуются характерные буро-черные кристаллы гематина. Последние часто склеиваются в более крупные включения. Кроме того, следует отметить, что внутриклеточные вакуоли с гемоглобином хорошо окрашиваются многими гистологическими красителями, включая железный гематоксилин, азокармин и эозин. Поступление крови в кишечник в значительных количествах происходит обычно не ранее середины периода роста, и к этому периоду относится и приводимое ниже описание пищеварительных клеток. Приблизительно с 4—5-го дня питания, по достижении веса 50—150 мг у Hyalomma asiaticum и 30—40 мг у Ixodes ricinus, в глыбовидных или булавовидных пищеварительных клетках появляются мелкие и средние белковые в'акуоли (диаметром 1—2 мк), дающие положительную реакцию на гемоглобин. Вначале они немногочисленны и локализуются преимущественно в краевой цитоплазматической зоне, но на более поздних стадиях могут заполнять всю апикальную половину клетки (рис. 323). Специальными методами окраски на липоиды в цитоплазме выявляются, правда, еще немногочисленные жировые вакуоли. Внешне пищеварительные клетки питающихся клещей во многом сходны с секреторными. Оба типа клеток претерпевают сходный процесс дифференцировки из резервных и на конечных этапах развития приобретают булавовидную или столбчатую форму. Пищеварительные клетки отличаются более крупными размерами, достигая у Hyalomma asiaticum величины 90—130x20—40 мк и у Ixodes ricinus — 100—140x30—50 мк.
Ядра располагаются ближе к центру клетки и достигают диаметра 20—30 мк. Базальная половина, как и у секреторных клеток, более узкая и заполнена пиронинофильными волокнистыми структурами. В апикальной цитоплазматической зоне отсутствуют крупные прозрачные вакуоли и ШИК-положительная зернистость.
По мере заполнения клетки гемоглобином более мелкие вакуоли, по-видимому, сливаются между собой, так как в цитоплазме появляются пищевые включения диаметром 3—5 мк. Появление кристалликов гема- тина обычно наблюдается лишь в клетках, загруженных пищевыми вакуолями и, вероятно, прекративших захватывание новых порций белка. Первоначально гранулы гематина немногочисленны, но по мере исчезновения пищевых вакуолей они в значительной степени загружают апикальные концы клеток и придают им характерную буро-черную или коричневатую окраску.
Пищеварительные клетки часто отторгаются в полость кишечника и плавают в ее содержимом в виде крупных сфер с ядрами и цитоплазмой, забитой различного рода включениями (рис. 314). В свободных клетках наблюдается нормальное продолжение процессов внутриклеточного переваривания гемоглобина, сопровождающееся исчезновением пищевых вакуолей и загрузкой цитоплазмы гематином. С окончанием усвоения содержимого пищевых вакуолей загруженные гематином свободные клетки разрушаются и их содержимое поступает в полость кишечника. Сходные процессы дегенерации наблюдаются и в пищеварительных клетках, связанных со стенкой кишки, по окончании в них цикла внутриклеточного пищеварения.
В первую половину периода роста среди гипертрофированных клеток преобладают секреторные, но постепенно количество пищеварительных клеток увеличивается и к началу периода растяжения они доминируют над остальными клеточными элементами. Тесно соприкасаясь своими дистальными концами, они, как и в случае с секреторными клетками на предшествующей фазе, образуют сплошной клеточный слой над вклинивающимися между их основаниями резервными клетками (рис. 323).
В период растяжения в связи с быстрым поступлением в кишечник большой массы крови хозяина происходит очень сильное растягивание стенок (рис. 312 и 324). Большая часть пищеварительных и секреторных клеток в конце питания разрушается или отторгается в просвет, а в стенках кишечника остается тонкий слой сильно уплощенных недифференцированных клеток высотой от 3 до 10 мк. Ядра также заметно уплощаются и на срезах имеют овальные или почти веретеновидные профили размером 20—30x4—8 мк. Границы между клетками на этой стадии становятся неразличимыми (рис. 324), а складки базальной мембраны и мускулатуры стенок полностью расправляются. Перед отпадением кишечник напитавшегося клеща находится в состоянии максимального растяжения и его стенки имеют вид тонкой мембрановидной оболочки, окружающей огромную массу непереваренной крови.
У отпавших клещей степень растяжения отростков кишечника несколько уменьшается за счет удаления из пищи излишней воды. Одновременно уже в первые сутки по отпадении отдельные растянутые клетки начинают увеличиваться в размерах и выдаются в просвет кишечника, приобретая сперва форму выпуклого свода, а затем обычную для гипертрофированных клеток форму дубинки или булавы (рис. 325, 326). По существу, этот процесс ничем не отличается от дифференцировки специализированных клеток из резервных во время питания и первыми его признаками по-прежнему служат появление в цитоплазме волокнистых элементов, богатых РНК, и рост ядер. У напитавшихся клещей дифференцировка резервных клеток идет в направлении пищеварительных, тогда как секреторные становятся сравнительно немногочисленными. В первые 1—2 дня по отпадении еще можно наблюдать отторжение отдельных, преимущественно секреторных клеток в просвет. Однако по мере исчезновения клеток этого типа процессы отторжения полностью прекращаются. Пищеварительные клетки напитавшихся клещей постоянно сохраняют связь со стенкой кишечника.
Образование новой «генерации» пищеварительных клеток связано с продолжением размножения недифференцированных клеток, среди которых в первые дни по отпадении наблюдаются многочисленные митотические деления. В результате уже к концу 2—3-х суток по отпадении стенка кишечника вновь образована сплошным слоем высоких пищеварительных клеток, между основаниями которых вклиниваются более мелкие резервные. Они сильно сжаты с боков пищеварительными клетками и приобретают клиновидную форму с широким основанием и заостренной вершиной (рис. 315 и 327). По-видимому, большая часть резервных клеток по окончании питания и во время яйцекладки находится на различных стадиях дифференцировки в пищеварительные клетки, так как их цитоплазма богата волокнистыми пиронинофильными элементами, а ядра увеличиваются в размерах. Некоторые исследователи (Till, 1961; Chinery, 1964) на основании высокой базофилии цитоплазмы выделяют их в самостоятельный тип железистых или печеночных клеток, но нам это кажется неоправданным. Такие же клетки, но в меньшем количестве встречаются и во время питания, причем можно проследить все этапы их превращения в специализированные пищеварительные или секреторные. Необходимо отметить, что после окончания яйцекладки у погибающих самок в стенках кишечника остается еще значительное количество не израсходованных в процессе пищеварения резервных клеток (рис. 329). К началу яйцекладки пищеварительные клетки необычайно увеличиваются, достигая высоты 400—500 мк при ширине 10—150 мк, т. е. в 40—50 раз превышают линейные размеры резервных клеток голодной особи. Ядра округлые или, реже, неправильной формы и достигают диаметра 20—35 мк, но часто не видны из-за многочисленных пищевых включений. Последние в виде средних и крупных вакуолей с гемоглобином крови хозяина имеют округлую или многоугольную форму на срезах и достигают в поперечнике 5—20 мк. Они заполняют всю цитоплазму, включая и апикальные концы клеток. Кроме белковых включений, на срезах, изготовленных на замораживающем микротоме и окрашенных Суданом черным, выявляется большое количество жировых вакуолей в виде капель различного размера (от 1 до 30— 40 мк).
После заполнения цитоплазмы белковыми включениями между ними появляются сперва одиночные, а затем в массе кристаллы гематина. Постепенно они заполняют всю апикальную половину клетки, после чего она разрушается, а гематин поступает в полость кишечника. Процессы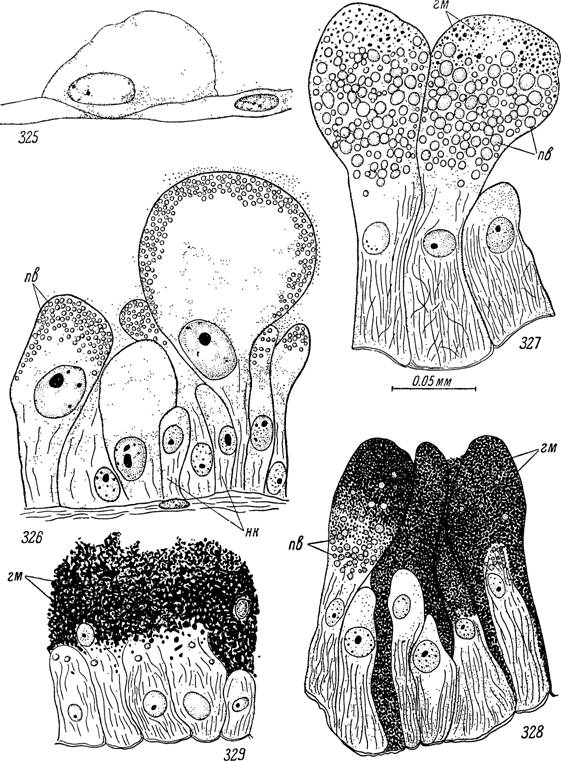
Рис. 325—329. Пищеварительные клетки отпавшей напитавшейся самки Ixodes
ricinus.
325 — начало дифференцировки пищеварительной клетки в растянутом участке стенки кишечника; гипертрофированные пищеварительные клетки: 326 — в первые дни по отпадении с хозяина; 327 — перед началом яйцекладки, 328 — в конце яйцекладки и 329 — у закончившей откладку яиц и погибающей особи.
пв — пищевые включения; гм — гематин; пк — недифференцированные клетки.пищеварения в отдельных клетках, даже расположенных поблизости друг от друга, не синхронизированы, так что одновременно встречаются как клетки, содержащие только пищевые вакуоли, так и полностью загруженные гематином (рис. 328).
Гипертрофированные пищеварительные клетки к началу яйцекладки поглощают всю кровь из полости кишечника. В результате просвет последнего сильно сужается, тогда как стенки дивертикулов необычайно утолщаются и в них концентрируются основные запасы питательных веществ, полученных во время питания. У заканчивающих откладку яиц самок большая часть пищеварительных клеток загружена гематином и подвергается разрушению. При этом полость кишки вновь несколько увеличивается и бывает заполнена буро-черной кашеобразной массой кристаллов гематина. Общий объем кишечника на этой стадии заметно уменьшается за счет частичного спадения стенок дивертикулов и последние свободно лежат в полости тела.
Самцы. Клеточный состав средней кишки у самцов тот же, что и у самок. У большинства непитающихся самцов рода Ixodes цикл изменений в средней кишке связан с постепенным расходованием пищевых резервов, накопленных на нимфальной фазе.
Процессы пищеварения у самцов с облигатным кровососанием рассмотрены на примере Hyalomma asiaticum и другого близкого вида — Н. plumbeum. У только что перелинявших самцов в клетках кишечника содержится значительное количество пищевых включений в виде округлых гранул различного размера (рис. 330). Постепенно запасные питательные вещества исчезают и у клеща, голодавшего длительное время, остаются единичные пищевые гранулы, а клетки уплощаются (рис. 331). Границы между отдельными клетками видны плохо, что вообще характерно для представителей этого рода.
После прикрепления к хозяину начинается интенсивный рост секреторных клеток в просвет кишечника, предшествующий началу кровососания (рис. 332). При поступлении крови отдельные клетки отрываются от стенок и перемешиваются с нею. Они бывают обычно сильно вакуолизированы и содержат зернистость полисахаридной природы (рис. 333). Одновременно начинается быстрое накопление пищевой зернистости в пищеварительных клетках. При ее переваривании, в отличие от самок, образуется меньшее количество гематина (рис. 334). В описанном состоянии кишечник самца находится до его гибели. Образование новых пищеварительных клеток непрерывно происходит за счет деления недифференцированных эпителиальных. Значительного растяжения стенок или образования «крипт» не отмечено.
Личинки и нимфы. Личинки Ixodes ricinus питаются 2—3 суток. Клетки кишечника голодной стадии очень мелкие и заметно уплощены. В период кровососания размеры их несколько увеличиваются. Рост их, однако, незначителен, и образующиеся глыбковидные клетки лишь в 2—4 раза выше нормальных. Этих размеров они обычно достигают через 24—36 час. кровососания. В боковом отростке на поперечном срезе бывает видно не больше 4—6 подобных клеток. Содержимое их сильно вакуолизировано.
За 10—12 час. до отпадения наступает быстрое растяжение стенок кишечника кровью и отрыв части пищеварительных клеток в просвет. У напитавшейся личинки стенки кишечника очень тонкие и состоят из немногих растянутых клеток. Накопления пищевых включений в период кровососания не отмечено. После отпадения среди оставшихся клеток наблюдаются митотические деления. Часть образующихся клеток вырастает в просвет и загружается пищевыми включениями, другие остаются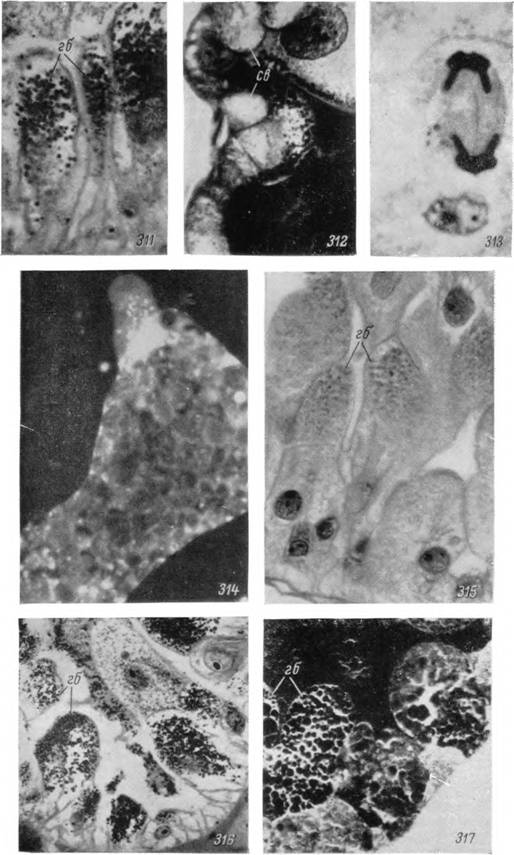
Рис. 311—317. Клетки средней кишки самки (311—315) и нимфы (316, 317) Hyalomma asiaticum на стадии питания.
311 — включения гемоглобина в пищеварительных клетках; 312 — участок стенки бокового отростка, заполненного кровью; 313 — митотическое деление недифференцированной клетки; 314 — отторгнутая в полость кишечника пищеварительная клетка; 315 — пищеварительные клетки напитавшейся особи; 316 — участок стенки бокового отростка в середине питания; 317 — то же у недавно напитавшейся особи. Микрофотографии: 314 — Х900, люминесцентный микроскоп после подкраски акридиновым оранжевым; 311, 312, 315— Х600; 313— Х2000; 316, 317 — Х4СЮ; окраска железным гематоксилином-эозином.
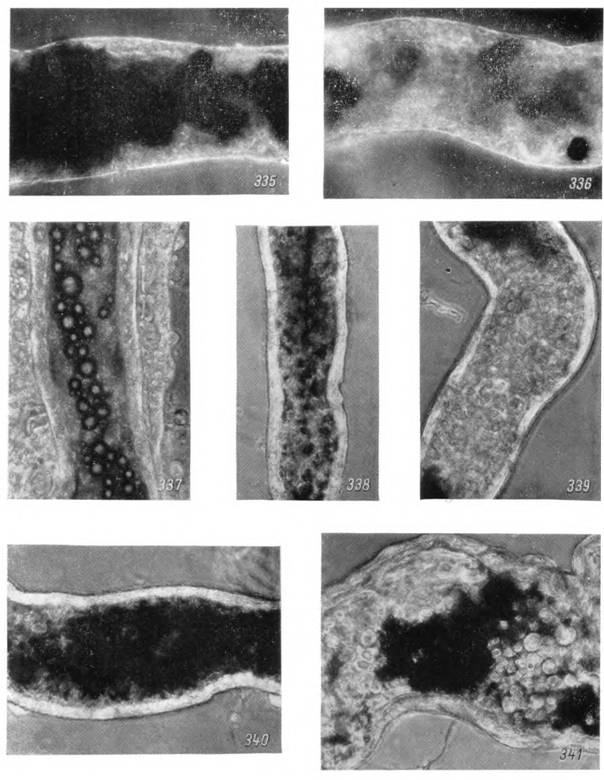
Рис. 335—341. Мальпигиевы сосуды Ornithodoros papillipes (335, 336) и Hyalomma asiaticum (337—341) на разных стадиях жизненного цикла.
335 — загруженный участок сосуда длительно голодавшей самки и 336 — участок сосуда вскоре после кровососания; 337 — нимфа на стадии линьки; 338 — голодная самка; 339 — самка в начале питания; 340 — в середине и 341 — в конце питания. Прижизненно в фазовом контрасте, Х200.в недифференцированном состоянии и позднее образуют кишечный эпителий нимфы.
Для личинок Hyalomma asiaticum, Rhipicephalus turanicus и Dermacen- tor pictus характерно накопление крупных пищевых включений еще в пе-
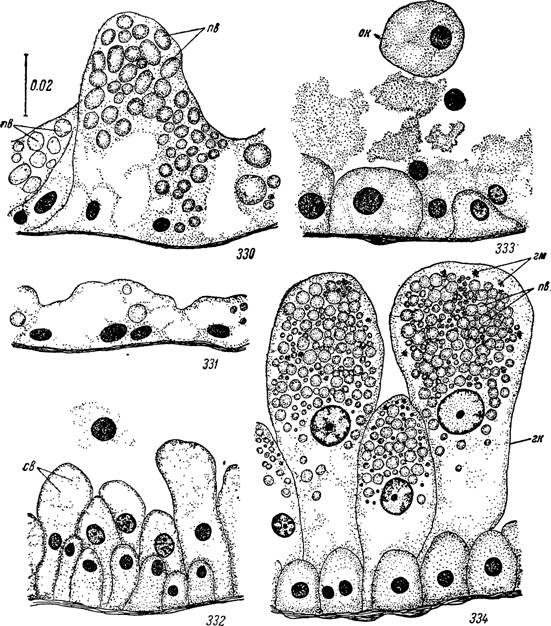
Рис. 330—334. Изменения кишечного эпителия у самца Hyalomma plumbeum во
время питания.
330 — сразу после линьки; 331 — на стадии активности; 332 — на 1-е сутки прикрепления к хозяину; 333 — на 2-е сутки и 334 — на 12-е сутки.
св — вакуоли с секретом; пв — пищевые включения; on — оторвавшиеся в полость кишечника пищеварительные клетки; гп — гипертрофированные пищеварительные клетки;
гм — гематин.
риод кровососания. Большая быстрота пищеварения у них связана с коротким периодом превращения в нимф.
У нимф пищеварение протекает в основном сходно с самками. Кишечник только что перелинявшей нимфы содержит много запасных питательных веществ в сохранившихся пищеварительных клетках личинки. В то же время пищеварительные клетки нимфальной фазы еще не обособились. У голодной нападающей нимфы клетки с пищевыми включениями почти полностью израсходованы. Пищеварительные клетки образуются в период кровососания и принимают глыбовидную форму. Содержимое их сильно вакуолизировано и многие из них отрываются в полость кишечника. Перед отпадением происходит быстрое растяжение стенок. У видов с быстрой линькой на взрослые фазы (Hyalomma, Rhipicephalus, Dermacen- tor) образование пищевых включений происходит уже в период кровососа- ния (рис. 316, 317), у видов с длительным периодом линьки, как Ixodes ricinus, — лишь после отпадения. Роет кишечника происходит за счет размножения недифференцированных резервных клеток. Митотические деления последних наблюдаются с начала периода роста и продолжаются всю первую половину линьки.
К концу стадии линьки между базальной мембраной и гипертрофированными пищеварительными клетками располагаются мелкие недифференцированные клетки — источник будущих секреторных и пищеварительных клеток половозрелой фазы.
Еще по теме ЦИТОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ПИЩЕВАРЕНИЯ:
- Общая характеристика работы
- Иксодовые клещи.
- ЦИТОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ПИЩЕВАРЕНИЯ
- ЗАКЛЮЧЕНИЕ
- Введение
- ДИАГНОСТИКА
- РАК ЖЕЛУДКА
- Заключение
- ВАЖНЕЙШИЕ КАТЕГОРИИ ПРИСПОСОБИТЕЛЬНЫХ И КОМПЕНСАТОРНЫХ АКТОВ B ФИЗИОЛОГИИ И ПАТОЛОГИИ
- Клинико-анатомические формы воспаления. Классификация. Терминология
- ОСНОВНЫЕ ЭТАПЫ РАЗВИТИЯ ПАТОЛОГИЧЕСКОЙАНАТОМИИ ВУКРАИНЕ
- ПРОФИЛАКТИКА ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ