СЛОЖНЫЕ ФОРМЫ ПОВЕДЕНИЯ И ИХ ПРОИСХОЖДЕНИЕ
Этологическая терминология, основанная на представлениях о неделимости поведения и о невозможности его нейрофизиологического, нейрохимического и структурного анализа, резко отличается от физиологической, используемой в учении о высшей нервной деятельности и нейрофизиологии.
Однако при изучении сложного поведения организма, складывающегося из врожденных и приобретенных элементов, как физиологи, так и этологи обращают внимание на одни и те же явления, обозначая их разными терминами.
В таблице 3 сделано некоторое сопоставление терминов, применяемых в физиологии высшей нервной деятельности, и терминов, применяемых этологами. Разумеется, некоторые из них приводятся весьма условно, но они все же позволяют ориентироваться хотя бы в тех аспектах, в которых проводится объективное изу іение поведения животных.
Таблица 3
Некоторые терминологические сопоставления из области высшей нервной деятельности и современной этологии
| Термины, приня'ые в физиологии 1 высшей нервні й деятельности | Термины этологические |
| Доминанта Адекватный раздражитель «Натуральный рефлекс» Безусловные рефлексы Натуральные условные рефлексы Искусственные условные рефлексы Уровень возбуждения, тонус нервной системы Обратная афферентация | Аппетенция Врожденный реализующий механизм Запечатлевание (импринтинг) Врожденное поведение, ин стинкт Облигатное обучение Факультативное обучение Зарядка центров Реафференция |
Многие расхождения между отдельными исследователями носят часто терминологический характер. Это касается, например, вопросов обучения и образования условных рефлексов, образования прочных условных рефлексов и явлений так называемого запечатлевания, проявлений доминанты и аппетенции, не говоря уже о терминах условнорефлекторного — «разумного» и безусловнорефлекторного — «инстинктивного» поведения.
Все эти различия в терминах не имеют в своей основе глубокого расхождения в понимании явлений, и это сильно затрудняет сопоставление фактов, полученных даже в близких методических условиях. Другое дело — глубокие методологические расхождения, касающиеся проблемы так называемой «реакции в пустоте» (Lecrlaiifreaktion), т. е. реакции, не вызванной факторами, действующими внутри или вне организма. Эта позиция многих этологов (Lorenz, 1937, 1957, 1960; Tinbergen, 1955; Eibl-Eiblsfeldt, 1958) не может быть ие признана фина- листской и здесь какие бы то пи было терминологические сопоставления, конечно, совершенно бесполезны.Современное представление о рефлекторной деятельности, изучаемой на уровне целого организма, включает детальное изучение как форм двигательной активности и ее чередования (поведение), так и ряда вегетативных процессов, которые вместе с отдельными формами поведения составляют целостные рефлекторные акты.
Уже сразу после рождения или вылупления из яйца организм способен реагировать на определенный комплекс раздражителей. Так, зрелые искусственно вынутые из яйцевых оболочек личинки жука тополевого листогры- за (Metasoma populi) задерживаются на листьях кормовых растений — тополя, ольхи и сразу покидают листья некормовых — например, березы или жасмина. Однако и на этих некормовых растениях они задерживаются дольше, чем на искусственных предметах, например бумаге, кинопленке и т. п. (Уждавини. 1957) (рис. 7). Следовательно, существует врожденная избирательная способность (врожденное узнавание и различение) дифференцировать непищевые и пищевые объекты. Влажная поверхность бумаги привлекает этих личинок больше, чем сухая. Значит, существует и определенная градация элементов этой дифференцировки от
более общих к более частным свойствам анализируемого субстрата.
Эти врожденные элементы «узнавания» дополняются образованием многочисленных условных рефлексов (временных связей). Значительный материал в этом направлении собран в лаборатории AV.
Е. Лобашова на пчелах и шелковичных червях.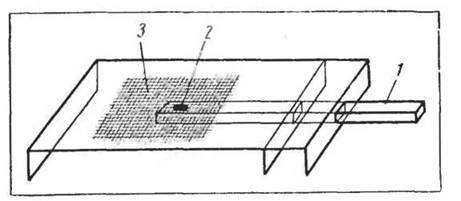
Рис. 7. Схема установки для изучения передвижения личинок но субстрату (по Уждавини, 1958): 1 — стержень для помещения личинки и субстрата, 2 — углубление стержня, 3 —■ миллиметровая сетка на поверхности столика
Обширный материал, свидетельствующий о значении слияния врожденного и приобретенного поведения у птиц, собран Н. AV. Промптовым (1956). Он показал, что поведение птиц н многие черты их пения в период размножения зависят не только от врожденных особенностей организма (видовые характеристики), но и от условий выкармливания своими или другими видами птиц. Особенно ярко выражены в этом отношении голосовые реакции птиц, выращенных родителями другого вида. Эти реакции являются довольно прочными, но могут изменяться на протяжении индивидуальной жизни.
У млекопитающих весьма сложные формы поведения можно наблюдать уже сразу после рождения до осуществления какого-либо контакта с факторами внешней среды. Наиболее общей реакцией, существующей практически у всех видов млекопитающих и вызывающей не только приближение новорожденного, но и пищевую реакцию, является положительная реакция на теплую поверхность.
Особенно важно отметить специфическое значение определенных факторов среды для появления отдельных 68 и притом специализированных форм поведения при экспозиции животных па разных стадиях онтогенеза. В этом случае — вне всякого образования условного рефлекса — возможно проявление чисто безусловного рефлекса, и такой адекватный раздражитель является в собственном смысле слова реализующим механизмом этологов (ange- borene auslosende Mechanismus). Так, например, у щенков, содержавшихся от рождения и до месячного возраста на молочном рационе, свойственная хищным положительная реакция на мясо появляется только на 20—21-й день после рождения, хотя мяса эти животные никогда до этого не если (рис. 8).
Другим более сложным примером может служить игровая деятельность у золотистого хомяка. Появление этой формы врожденной деятельности на определенном этапе онтогенеза не зависит от факторов внешней среды, но в то же время может усиливаться при образовании соответствующих условных рефлексов и усиливаться и тормозиться внешними температурными влияниями (рис. 9, 10). Таким образом, хотя собственно «реализатором» этой формы врожденной деятельности является только партнер по игре, предшествующие раздражения (температурные), условные рефлексы на обстановку (систематическое отсаживание в клетку для игры), время опыта несомненно имеют в этом случае немаловажное значение. Однако здесь имеет место и определенная «зарядка» центров, которая как бы отражает накопленный нервными центрами потенциал. Если представление об аккумуляции энергии центрами является в данном случае только гипотетическим н ничего не объясняющим, то само состояние повышенной возбудимости и работоспособности выполняющих эту форму деятельности центральных и периферических аппаратов не вызывает никакого сомнения.
Если характеризовать с этой стороны врожденные формы деятельности, то на основании уже многих исследований (Попугаева, 1959, 1960; Рахимов, 1958; Ужда- вини, 1955, 1958; Щеглова, 1958, 1958а; Шепелева, 1960, 1963), можно утверждать, что существуют определенные сроки постнатального развития, когда эти реакции вызываются ничтожными по силе (подпороговыми) внешними раздражениями из внешней среды или даже возникают «спонтанно». В последнем случае нет никакой возмож-
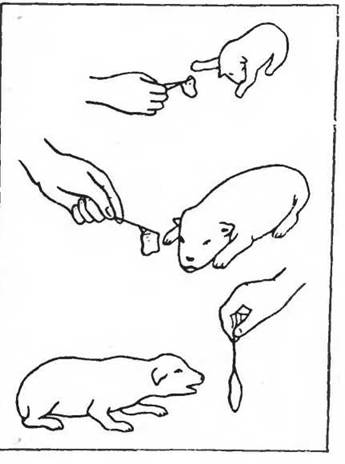
Рис. 8. Двигательная реакция на мясо у щенков домашней собаки (по Ужда- винн, 1958); вверху—1-й день после рождения, в середине— 2-й день, внизу — 21-й день
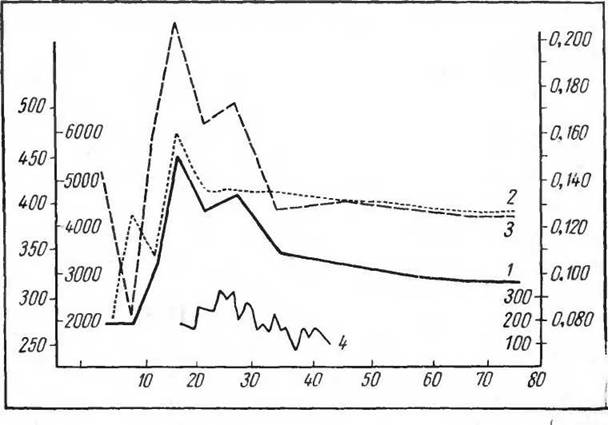
Рис. 9. Изменение некоторых функций в онтогенезе у золотистых хомяков (по Понугаевой, 1968):
1 — газообмен, 2 — пульс, 3 — кислородный пульс, 4 — игровая деятельность.
По оси абсцисс — возраст (дни); по левой оси ординат слева — частота пульса, справа — газообмен (О2 мл'кг ч); по правой оси ординат слева — длительность игровой деятельности (сек), справа — кислородный пульс (О2 мл'кг веса на 1 удар пульса)ности установить характер «внешнего раздражения, вызывающего сформированную к этому периоду развития рефлекторную реакцию. Однако речь идет, конечно, только о внешних стимулах, сравнительно легко доступных регистрации и изучению. Внутренняя стимуляция, включая собственно стимуляцию, возникающую в самом
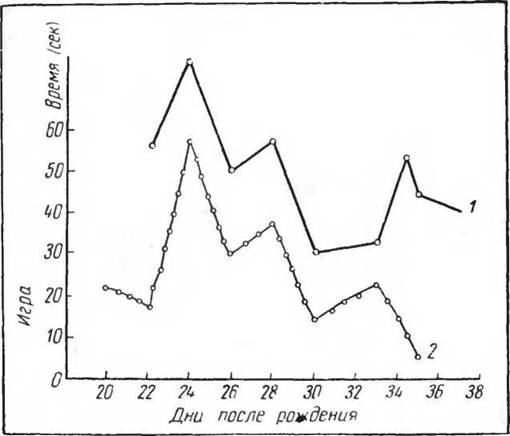
Рис. 10. Игровая деятельность золотистого хомяка (сек), (по Понугаевой, 1968) при содержании при разных температурах среды:
1 — 12° С, 2 — 25° С
нервно-мышечном приборе, остается вне поля зрения исследователя. Эта очень важная сторона вопроса до настоящего времени почти не изучена.
С этой точки зрения большой принципиальный интерес представляет положение Делла (Dell, 1958) о взаимоотношениях между анимальными и вегетативными функциями в организме. По Деллу, аппетентная стадия деятельности организма есть «превращение нужд тела в поведение»—(transformation of bodily needs into behaviour). Механизмы этого превращения выявляются в современной физиологии в виде стимулирующего действия на головной мозг восходящей части ретикулярной формации стволовой его части, гормональных сдвигов (кора надпочечников). Однако современные знания находятся еще на подступах к решению этой кардинальной проблемы.
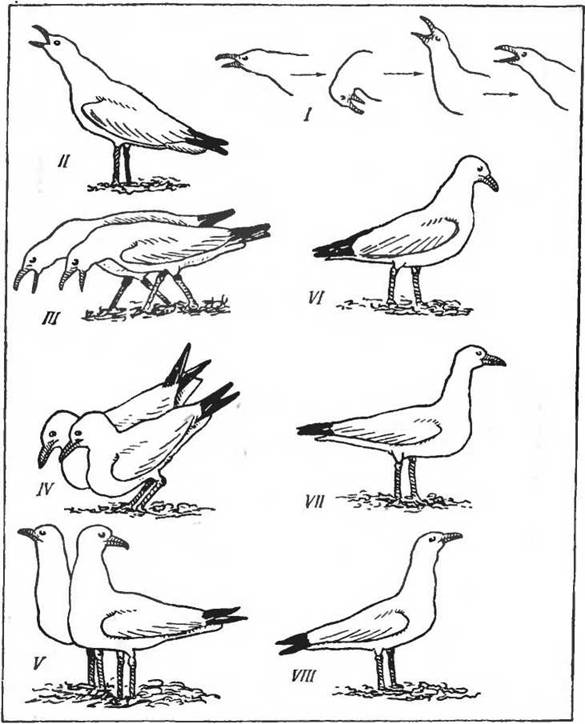
Рис. 11. Формы поведения у чаек (по Tinbergen, 1965). I — движения у сизой чайки при длительном призывном крике; II—V — церемония встречи у серебристой чайки (// — призывный крик, III — общий зов, IV — «давятся», V— отворачивают голову, смотрят в разные стороны); VI—VIII — стоячая лоза у серебристой чайки (VI — угроза, VII — неуверенность, VIII — страх)
Несмотря на то, что индивидуальные элементы поведения (благодаря образованию многочисленных условных рефлексов) могут различаться у разных особей одного и того же вида, существует четко выраженная
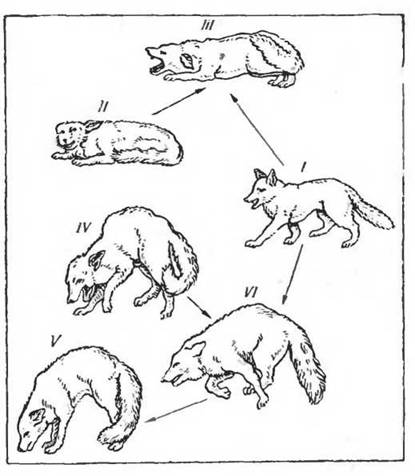
Рис.
12. Типичные формы поведения у красной лисицы (по Tembrock, 1964). I— нападение; II— пассивно-оборонительная реакция (тенденция к бегству); III — наложение состояний I и II (полное торможение двигательной активности); IV — оборонительное поведение против укуса сзади; V— наложение состояний І м IV (нападение с согнутой спиной); VI — ритуализированное поведение подчиненияспецифика в формах поведения, реакциях на определенные раздражители у разных, даже близких в систематическом отношении видов. Эта называемая этологами «морфология поведения» служит важным таксономическим признаком и представляет одну из наиболее характерных черт сложных форм поведения. На рисунках 11, 12 приведены примеры такого поведения у представителей птиц и млекопитающих. Изучение этих особенностей
поведения у отдельных популяций одного и того же вида представляет огромный интерес для решения экологофизиологических проблем.
Еще по теме СЛОЖНЫЕ ФОРМЫ ПОВЕДЕНИЯ И ИХ ПРОИСХОЖДЕНИЕ:
- Происхождение установки в автоматизированном индивидуальном поведении.
- 30.Сущность и формы отклоняющегося поведения личности.
- 41.врожд формы поведения.безус реф
- 31. Неактивные формы поведения животных: общая характеристика.
- 52. Сложные нарушения развития. Подходы к классификации. Психическое развитие при сложных нарушениях
- 21. Делинквентное поведение как форма отклоняющегося поведения личности.
- 1. Предмет и содержание дисциплины «Организационное поведение». Модели поведения человека в организации
- Параграф пятый. О сложных болезнях
- Параграф одиннадцатый. Сложные лекарства
- Образование условных рефлексов на сложное раздражение.
- Дискуссия! Является ли человеческийорганизм более сложным и совершенным, чем ПК?