Свойства селена, механизм транспорта и биохимических превращений в клетке
Se относится к IV группе элементов периодической таблицы и следует за серой. Химические свойства серы и селена сходны. В биологических макромолекулах селен находится в следующих формах: селенол (-SeH), селенид (HSe) и селенолы (RSe).
Селенит может поступать в клетку и восстанавливаться в селенид специфическими системами. Свободный селен, практически не ассимилируется организмами. Селениты (K2SeO3) и селенаты (K2SeO4) с разной степенью активности.В кислых почвах селен находится в виде селенита железа, мало растворим в воде и не ассимилируется организмами. В щелочных почвах селен находится в окисленной форме, ионы селена га растворимы, хорошо ассимилируются организмами и включаются в состав органических соединений в одной из трех форм:
—Н2С—Se—СН2— -- Н2С—S—Se—СН2— —Н2С—Se—Se- СН2 —
Возможны различные пути поступления селена в клетки микроорганизмов. Селенат и сульфа і могут поступать в клетку по одному пути. Селенат является конкурентным ингибитором энергоза висимого транспорта сульфата. Конкуренция между селенатом и сульфатом происходит на уровн< адсорбции.
На сульфат-селенатный антагонизм влияют три фактора:
• разные скорости адсорбции сульфата и селената;
• уменьшение адсорбции селената при увеличении экзогенного сульфата;
• конкуренция между двумя ионами за один фермент.
Кинетика поступления селената описывается уравнением Михаэлиса-Ментен с величиной Км = 1,43 мкМ.
В бактериях в аэробных условиях и в бактериях Е. coli и Clostridium в анаэробных условиях по казано существование разных систем транспорта селенита, сульфита и селената. В клетки дрож жей сульфат и селенит поступают по одной транспортной системе. Закономерности конкуренции между серой и селеном зависят от концентрации селенита в среде.
Метаболизм селена. В клетках микроорганизмов выявлена система восстановления селенита селенредуктазой, аналогичная системе ассимиляционной сульфатредукции, восстанавливающая селенит до элементарного селена. Восстановление селенита осуществляется с большей скоростью, чем селената. Восстановление селенита в Se2' может происходить при участии глутатиона и глута- тионредуктазы.
селенит
2НАДФН гНАДФ+GSH 2НАДФН гНАДФ+GSH
4GSH GSSG
H2SeO3

GSSeSG

GSSeH
GSH-редуктаза
H2Se
GSH-редуктаза
селеноди-
глутатион
селенпер-
сульфид
При взаимодействии селенита с 2 молекулами восстановленного глутатиона (GSH) образуется селенодиглутатион (GS—Se—SG), который является ключевым продуктом в дальнейших путях превращения селена в клетке. Он может либо восстанавливаться при участии глутатионредуктазы до лабильного селенперсульфида (GSSeH) при избытке глутатиона, либо возможно каталитическое окисление селенодиглутатиона с высвобождением внутриклеточного селенита, что зависит от соотношения концентрации селенита и глутатиона в клетке. При низких концентрациях селенита протекает реакция каталитического окисления глутатиона. При высоких концентрациях селенита происходит образование селенистого водорода (H2Se). H2Se может фиксироваться на белках. Селенодиглутатион (GS—Se—SG) может взаимодействовать с другими тионами, особенно с сульфгид- рильными группами некоторых белков. Эти реакции объясняют токсичность больших доз селена для организмов.
Селенистый водород, одна из токсичных форм селена, может быть окислен до элементарного селена кислородом воздуха или подвергаться метилированию с образованием летучих соединений селена — (CHa)2Se, CH3SeH.
Селенистый водород H2Se может выделяться из клетки в анаэробных условиях, он может захватываться кислотолабильным белком типа альбумина и включаться в процесс синтеза селеносодержащих аминокислот, метионина и цистеина. Оба эти пути являются механизмом детоксикации селена клеткой.Селенометионин неспецифически включается в селенсодержащие белки и полипептиды. Селеноцистеин специфически включается в селензависимые ферменты электронтранспортной цепи, металлотионеины, входящие в состав тРНК, в состав ферментов, изменяющих насыщенность липидов и жирных кислот. Обнаруживаются также селенаналоги КоА и тиамина.
Токсическое действие высоких концентраций селена на организмы вызвано замещением селеном атомов серы в биологически значимых соединениях и вследствие этого изменением их функций. Замещение метионина на селенометионин может оказывать влияние на структуру белка, но не на его каталитические свойства. В противоположность селенометионину, включение селеноцистеиновых остатков в белки является высокоспецифичным. Замещение серы селеном в цистеине, который входит в состав окислительно-восстановительных ферментов, влияет на каталитические свойства этих ферментов. В клетках анаэробных или факультативных анаэробных бактерий функционирует ряд селензависимых ферментов (глицинредуктаза, формиат и другие дегидрогеназы, гидроксилазы и др.)
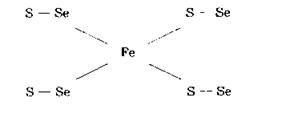
Селениты и селениды могут включаться в большую группу белков с образованием селенотри- сульфидных (R-S-Se-S-R), либо селеноперсульфидных (R-S-Se-H) мостиков. Селениды могут замещать сульфиды и в белках, содержащих железо.
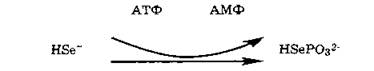
Включение селенида в белки происходит в несколько этапов. На первом этапе специфическая т РНК нагружается серином при участии серил — т РНК — синтетазы и образуется серил — т РНК.

Селенид (HSe) активируется селенофосфатсинтетазой и образуется селенофосфат (HSePO32).
На основе активированного селенида (селенофосфат) образуется селеноцистеин, что катализируется пиридоксальфосфатсодержащей селеноцистеинсинтазой.
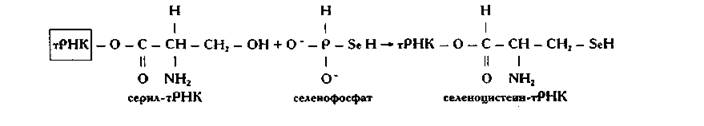
Селеноцистеин включается специфично в белки ферментов и кофакторов клетки. Существует ряд ферментов в клетках эукариот и прокариот, включающие селен в форме селеноцистеина.
4.3.
Еще по теме Свойства селена, механизм транспорта и биохимических превращений в клетке:
- Распределение селена, аккумулированного в клетке микроорганизмов
- Механизмы белкового транспорта
- Основные физиолого-биохимические свойства штаммов-продуцентов рибофлавина
- Основные общие механизмы повреждения клетки
- Определение экспрессии Толл-подобных рецепторов (TLRs) на клетках крови, клетках кожи и клетках эпителия слизистой зева.
- Механизмы токсического повреждения систем энергообеспечения клетки
- Превращение и улучшение.
- Накопление селена в биомассе
- Казарьян, Константин Александрович. Биохимические и иммунологические свойства Белков семейства Rpf — факторов роста [Электронный ресурс] Micrococcus Iuteus и Mycobacterium tuberculosis : Дис. ... канд. Биол. наук : 03.00.04 .-М.: РГБ, 2003, 2003
- Механизм влияния сочетанного применения эмоксипина и димефосфона в базисной терапии больных миомой матки на клиникобиохимические показатели, реакции ПОЛ и эндогенную интоксикацию