Генетика и селекция S. avermitilis
К настоящему времени установлена практически полная последовательность генома линейной хромосомы S. avermitilis. Геном содержит 9 025 608 нуклеотидов (среднее содержание суммы оснований гуанина и цитозина 70,7 %).
Кластеры генов, ответственные за биосинтез вторичных метаболитов, составляют 6,6 % генома. Кластер генов биосинтеза авермектинов состоит из 18 генов и содержит 80 192 пар нуклеотидных оснований (около 1,8 % генома). Для всех 18 генов дедуктивно (на основе сравнительных данных по гомологии с генами других организмов) рассчитаны их белковые продукты (и последовательности аминокислот в них) и возможные функции для 17 белков: 16 ферментов и одного регуляторного белкаАое R. Один из генов, входящих в кластер, кодирует белок, не участвующий в синтезе авермектинов.Выделенный из почвы (в Японии) штамм S. avermitilis МА-4680 исходно продуцировал 9 мг/л авермектинов. После физического мутагенеза (УФ-облучение) споровой суспензии исходного штамма и подбора ферментационной среды и условий культивирования был отобран мутант (АТСС 31271), образующий до 500 мг/л авермектинов, в том числе около 160 мг/л компонента В1. Этот мутант стал исходным для дальнейшей селекции более продуктивных штаммов.
В качестве типичной процедуры селекции, используемой для получения новых штаммов, можно привести следующие данные. Авторы использовали отбор как спонтанных мутантов, так и индуцированных путем УФ-облучения и химического мутагенеза (обработка метилметаносульфонатом и нитрозокислотой). Эффективность мутагенеза контролировали по изменению 4 фенотипических признаков:
• морфологические: образование воздушного мицелия (ату) или спор (spo);
• образование пигмента меланина (mel);
• образование авермектинов (ave);
• устойчивость и стрептомицину (str11).
Исходным для селекции служил штамм АТСС 31271. Спонтанные мутанты этого штамма отличались значительной вариабельностью фенотипических признаков.
Среди не образующих авермектины мутантов (аое~) 92 % клонов имели фенотип mel~, 78 % клонов — ату~, 32 % клонов — spo~ и 47 % клонов — strR . С фенотипом ату+/spo*/mel+ 8,7 % клонов не продуцировали авермектины, и только 0,22 % клонов синтезировали компонент В1 в количестве большем, чем штамм АТСС 31271. Один из них, штамм 267/2Н, продуцировал до 230 мг/л В1 (против 162 мг/л у исходного штамм). Путем мутагенеза из штамма 267/2Н был получен морфологически стабильный мутант с фенотипом spo+/mel+/str^, образующий до 466 мг/л В1.Наиболее высокая продукция авермектинового комплекса, о которой сообщалось в открытых публикациях, составляет от 1800 до 4000 мг/л. В комплексе, синтезируемом штаммом S. avermitilis НИЦБ-132 (Россия), содержание компонентов фракции В1 достигает 42 % при общей продукции авермектинов в пределах 3500-4000 мг/л.
Процедура получения штамма НИЦБ-132 была основана на ступенчатой селекции мутантов, устойчивых к антибиотикам и аналогам аминокислот (табл. 1). Мутация устойчивости к хлорамфениколу влияет на регуляцию механизма, связанного, очевидно, с повышением активности проте- инкиназ(ы), контролирующих экспрессию позитивного транскрипционного регулятора Ave R. Мутации устойчивости к аналогам изолейцина и треонина ведут к увеличению внутриклеточного пула МКВ (см. рис. 2), предшественника стартовой единицы авермектинов фракции «а», и, как следствие, к повышению доли В1а во фракции В1 (а+Ь).
С целью получения штаммов, образующих только компоненты В1а и В2а, предварительно были отселекционированы путем УФ-облучения и химического мутагенеза штаммы, не образующие компоненты фракции В, и штаммы с поврежденной 5-0-метилтрансферазой, не образующие компоненты фракции А. На следующем этапе селекции были использованы методы генной инженерии: перекрестный перенос генов в протопласты этих штаммов и самослияние протопластов, в результате чего был отобран рекомбинантный штамм К2032, синтезирующий компоненты В1а и В2а в количестве 0,2 г/л при содержании В1а 65 %.
Из этого штамма после мутагенеза был получен штамм К2056, образующий 1,0 г/л В1а и В2а при содержании В1а 55 % . В дальнейшем в хромосому штамма К2038 с помощью плазмиды интродуцировали фрагмент ДНК, содержащий участок с мутацией гена ave С, и отобрали штамм, синтезирующий только авермектин В2а, из которого путем химической трансформации можно получать 22, 23-диоксиавермектин В1а.На основе природных авермектинов можно получать новые производные не только химической, но и биотрансформацией. Эту возможность представляет, прежде всего, субстратная неспецифич- ность начальной реакции в биосинтезе лактона — конденсация стартовой единицы к метилмало- нил-КоА (см. рис.2). Оказалось, что в качестве стартовой единицы могут выступать, помимо изо-
Табл. 1. Селекция S . avermitilis
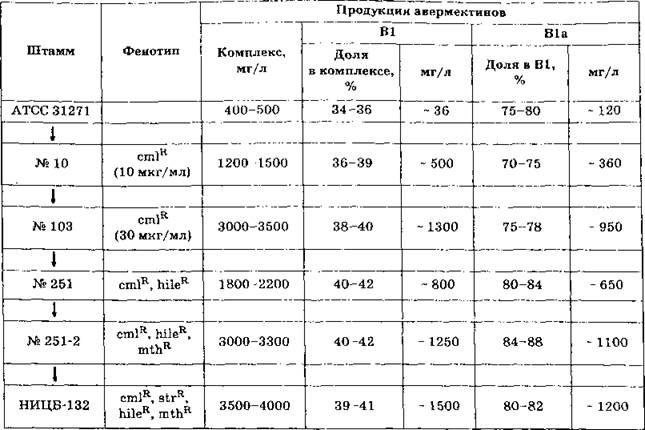
Примечание: cml R — устойчивость к хлорамфениколу; hileR — устойчивость к гидроксамату изолейцина, mthR — устой чивость к О-метилтреонину
и 2-метил-масляной кислот, и другие разветвленные жирные кислоты, содержащие 5-8 атомов углерода. Это было продемонстрировано в культуре штамма с поврежденной дегидрогеназой разветвленных 2-кетокислот (Bkd; см. рис.2), неспособного синтезировать изо- и 2-метил-масляную кислоту. В присутствии в культуре этого штамма других короткоцепочечных разветвленных жирных кислот синтезировались авермектины, содержащие в позиции радикала R2 (см. рис. 1) соответствующие добавленные кислоты.
Одним из наиболее перспективных производных авермектина, полученных с использованием штамма bkd- является дорамектин (коммерческое наименование «Дектомакс»), Стартовой единицей при биосинтезе этого вещества является си-циклогексанкарбоксикислота (ЦГК). Селекция штамма, образующего ЦГК-В1 (дорамектин), может служить ярким примером тех возможностей современных методов генной инженерии и протеомики, которые позволяют получать продуценты новых «гибридных» форм биологически активных веществ.
ЦГК является частью антибиотика анзатринина А, синтезируемого S. collinus. Была сконструирована плазмида, содержащая 5 генов, кодирующих ферменты биосинтеза ЦГК, которую трансформировали в штамм S. avermitilis bkd~, ave D~. Образующаяся в клетках S. avermitilis ЦГК активируется до ЦГК-КоА, которая служит стартовой единицей и включается в состав компонентов фракций «1» и «2» с образованием ЦГК-В2 и ЦГК-В1 в соотношении 1,6:1. С помощью сайт-спеии- фического мутагенеза гена ave С была установлена роль отдельных аминокислот в составе Ave С (дегидратазы) в катализе образования двойной связи между атомами 22С и 23С в лактоне. Затем методом «РНК-полимеразной реакции» были синтезированы гены ave С с модифицированными кодонами, которые клонировали в хромосому нового штамма. В результате был отобран клон, образующий ЦГК-В2 и дорамектин в соотношении 1:2, что значительно повысило продуктивность ферментационного получения нового коммерчески важного препарата.
Теперь, когда геном S. avermitilis охарактеризован практически полностью и все более идентифицируются функции отдельных генов, создаются реальные предпосылки эффективно использо вать для получения мутантов, синтезирующих производные авермектинов с заданной композицией, методы геномики и протеомики.
Единственным регуляторным белком, позитивно влияющим на транскрипцию всех структурных генов, входящих в кластер генов биосинтеза авермектинов, является Ave R. Убедительным доказательством может служить тот факт, что увеличение активности 5-0-метилтрансферазы, сопровождаемое повышением общей продукции авермектинов, не изменяет соотношения фракций Г и А, которое остается постоянным (6:4).
Известно, что регуляторные белки обычно находятся под контролем протеинкиназ и проявляют свое регуляторное действие в фосфорилированной форме. Функционирует ли подобный контроль Ave R , — пока неизвестно. Однако была обнаружена большая активность протеинкиназ у cml- устойчивых мутантов S. avermitilis, отличающихся повышенной продукцией авермектинов (см. табл. 1), по сравнению с cznZ-чувствительным штаммом. В ферментационной культуре cznZ-устойчи- вого штамма в период перехода в стационарную фазу роста в клетках наиболее заметно повышается самофосфолирование аминокислот серина и/или треонина в 4 белках с молекулярной массой 45, 52, 62 и 85 кДа. Сравнение этих белков (по значениям молекулярных масс и pi) с серин/треонин- протеинкиназами (СТПК), идентифицированными у S. avermitilis, позволяет отнести белки 45, 52 и 62 кДа к СТПК, кодируемым генамиpkn 24, pkn 13 и pkn 12, соответственно. Белок 85 кДа не был идентифицирован, однако, предположительно, также является СТПК, зависимой от Са2+ и калмо- дулина. Какова роль обнаруженных протеинкиназ и их иерархия в цепи регуляторных реакций, определяющих интенсивность биосинтеза авермектинов, — предмет дальнейших исследований.
6.