2.3.7 Перекисное окисление фосфолипидов
в той или иной степени ответственно за повреждение мембран эндоплазматического ретикулума в клетках неоплазмы. Наряду с дезынтеграцией микросомальных мембран происходят существенные изменения состояния монооксигеназной ферментной системы и, в первую очередь, главного её компонента – цитохрома Р-450 (CYP), под наз-ванием которого числится фактически целая группа его изоферментов.
Как известно, эти ферменты участвуют в большом наборе реакций с множеством субстратов: нормальных клеточных компонентов, модификация которых составляет нормальный процесс их метаболизма, и чужеродных соединений, превращающихся в ряде случаев в продукты с патологическими свойствами, в том числе канцерогенными. Условные изображения 2 типов оксигеназных реакций с включением одного или обоих атомов О2 в субстрат (Бохински, 1987) показаны ниже:SH2 + DH2 + O2 → HS-OH + H2O + D;
SH2 + O2 → HO-S-OH,
где SH2 – донор электронов, а DH2 – органическое соединение в роли донора водорода.
Основные исследования по состоянию монооксигеназной ферментной сис-темы в опухолях проводились на перевиваемых гепатомах. Результаты свидетельствуют в пользу того, что в опухолях содержание CYP и b5 и их функци-ональная активность снижены по сравнению с гомологичной нормальной тканью. В них, как правило, нарушено и характерное для нормы соотношение изоформ CYP (Кобляков, Коляда, 1986). Так, в клетках медленнорастущей гепатомы Морриса 9618А уровень фермента CYP снижен на 80 %, а в клетках быстрорастущей гепатомы 3924А он фактически отсутствует (Massoti et al., 1989). Состояние монооксигеназной ферментной системы в опухолях человека освещено лишь в единичных работах. Например, было показано, что у 5-ти исследованных гепатобольных содержание CYP в опухолях было снижено примерно в 10 раз по сравнению с его уровнем в окружающей ткани (Sasaki et al., 1984). Из 16-ти находившихся под наблюдением онкобольных у 12-ти мета-болическая активность CYP2С19 была ниже, чем у здоровых испытуемых (Wiliams et al., 2000).
При активации NADPH-специфичного и аскорбатзависимого ПОЛ окис-ляются прежде всего ненасыщенные жирные кислоты из фосфолипидного окружения CYP, где расположены радикалинициирующие центры.
При этом фермент CYP, очевидно, инактивируется, но после восстановления состояния указанного его окружения он в принципе готов к повторному «включению». Зависимость активности CYP от фосфолипидов показана, в частности, на при-мере CYP1А2. Наиболее эффективными активаторами его были фосфатидил-холины с числом атомов углерода в углеводородной цепи 8-12 (Yun et al., 1997). В свою очередь, содержание CYP и липидов является лимитирующим фактором в активности переокисления липидов. Токсические продукты, образующиеся при NADPH-зависимом перекисном окислении микросомальных липидов, достаточно стабильны и способны диффундировать, т.е. могут вызывать ток-сический эффект на некотором расстоянии от места протекания липидной пероксигенации (Benedetti et al., 1979).Названные выше эффекты в основном известны давно, но сведения о них периодически уточняются и/или пополняются новыми фактами. Например, в недавнем обстоятельном обзоре (Арчаков и др., 1998) рассмотрены механизмы реакций окислительной самоинактивации и модификации CYP и некоторых других макромолекул. Подчёркнута важная особенность CYP: он не только генерирует Н2О2 и иные АФК, но и сам инактивируется под их действи-ем. Детальнее исследован механизм окислительной модификации CYP2В4 в монооксигеназной реконструированной системе, включающей также NADPH-специфичный флавопротеин и цитохром b5. Показано, что окислительное изменение CYP сопровождается деструкцией гема и модификацией апофермента. Кроме того, АФК участвуют в окислении SH-групп белка-фермента и в образовании карбонильных групп на его молекуле. Как полагают авторы, самоинактивация и модификация CYP и других ферментов под действием АФК в процессе катализа могут лежать в основе механизма удаления их эндогенными проте-азами. Вообще же, это связано с тем, что у окисленных белков нарушается пространственная структура, они утрачивают активность и становятся высокочувствительными к протеолизу (Dean et al., 1997).
В условиях гипероксии и повышенной пероксигенации, характерных для опухолевых клеток, указанные выше изменения в микросомах должны быть, очевидно, более выраженными.
К тому же, окислительный стресс оказывает негативное влияние на регуляцию транскрипции некоторых генов CYP. В частности, обработка клеток HepG2 человека или гепатомы Н4 крысы пероксидом водорода подавляет связывание ядерного фактора NF1 с его сайтом в ДНК, локализованном в проксимальном промоторе гена CYP1А1. Предположительно, NF1 обладает повышенной чувствительностью к окислительному стрессу, что и приводит к ингибированию активности промотора и соответственно подавлению экспрессии CYP1А1 (Morel, Barouki, 1998). Определённый вклад в инактивацию ферментов микросом может вносить и синтезируемый в клетках неоплазмы NO. Показано, в частности, что in vivo NO ингибирует CYP, и этот процесс, по мнению Хаценко (1998), реализуется по двум механизмам: прямое влияние на активность фермента путём связывания с его гемом и угнетение экспрессии гена CYP.С другой стороны, обращено внимание на риск развития ракового заболевания при сверхэкспрессии CYP, поскольку существует корреляция между индукцией его изоферментов и уровнем О2 – одним из ведущих факторов канцерогенеза (Paolini et al., 1996; см. также п. 2.3). И действительно, к настоящему времени накоплен весомый экспериментальный материал об индукторах суперсемейства CYP как, прежде всего, промоторов канцерогенеза. Многие вопросы, касающиеся механизма действия этих индукторов, подробно рассмотрены в работе Коблякова (1998). Из числа цитированных и обобщённых им литературных материалов и высказанных идей наиболее интересными, с точки зрения кислородно-перекисной модели канцерогенеза, представляются нам все эффекты индукторов CYP, опосредованные действием образующихся при функционировании данного фермента АФК.
Как принципиально важные, выделим: митогенное действие АФК (Burdon, 1992, 1995; Burdon et al., 1996; Bhunia et al., 1997 и др.) и активацию ими некоторых факторов транскрипции (Flohe et al., 1997; Renard et al., 1997; Mazi-ere et al., 1999; Турпаев, 2002 и др.); нарушение межклеточных коммуникаций кислородным стрессом (Hu Y.
et al., 1995; Lo et al., 1998; Wenk et al., 1999), вызываемое, предположительно, активацией АФК онкобелка c-src (Lee et al., 1996; Griendling et al., 2000), фосфорилированием последним белков межклеточных контактов, в частности коннексина-43, и выходом их из мембраны клетки (Yamasaki, Naus, 1996 и др.); участие АФК в высвобождении из ферритина ионов Fe, катализирующих образование различных АФК (Puntarulo, Cederbaum, 1996); активацию АФК выхода из мембраны свободной арахидоновой кислоты (Martinez, Moreno, 1996) и последующий её метаболизм на CYP с образованием высокореактивных О2-содержащих метаболитов и новых АФК.Опираясь на указанные выше работы и мнение о белке c-src как первичном объекте воздействия АФК (Devary et al., 1992; Lee et al., 1996), Кобляков (1998) высказал также свою точку зрения, отметив, что «митогенный эффект и блокирование межклеточных коммуникаций при действии АФК реализуются через общее звено посредством активации продукта гена c-src», причём «не исклю-чено существование некоего неизвестного рецептора АФК, который приводит в действие продукт гена c-src, запуская каскад киназных реакций». Поскольку конкретные механизмы активации обсуждаемых АФК-зависимых эффектов индукторов CYP пока неизвестны, то естественно ожидать получения здесь каких-то важных «связующих» фактов. Можно, например, проверить, причастны ли внутриклеточные АФК и продукты ПОЛ, образующиеся в избытке под влиянием промоторов, к хотя бы некоторым эффектам последних за счёт известной способности их активировать PLA2. Активация белка c-src проокси-дантами также может происходить косвенно, а пока, судя по публикациям последних лет, известно, что модуляция активности продукта гена c-src осуществляется в основном протеинкиназами, в частности, некоторыми рецептор-ными тирозинкиназами (см. п. 3.4.2).
Обсуждая роль CYP как промотора канцерогенеза, важно знать также, ка-кие из образующихся в клетке продуктов способствуют развитию при участии CYP ракового заболевания. В этом отношении заслуживают внимания следую-щие научные материалы. В микросомах печени человека был обнаружен фермент, превращающий простагландин-эндопероксид в гидроксигептадекатриеновую кислоту и MDA.
Этот процесс не зависел от CYP-редуктазы, но связан с CYP. Активности очищенных CYP к указанному субстрату (50 мкМ) варьировали, и по этому показателю некоторые его изоформы располагались в последовательности: 3А4 >> 2Е1 > 1А2. Авторы данной работы (Plastaras et al., 2000) допускают, что при коэкспрессии COX-2 и изоформ CYP в процессе канцеро-генеза может возникать нестабильность генома, обусловленная образованием MDA – эндогенного мутагена. Возникновение такой канцерогенной ситуации в дополнение к создаваемой недостаточностью митохондриального дыхания вполне реально, если учесть избыточные синтез и активность COX-2 при канцерогенезе (см. п. 2.1.11).Процесс деградации микросомальных мембран естественно связать с высвобождением интегрированных с ними рибосом. Это объяснило бы известный феномен: в цитоплазме опухолевых клеток резко уменьшена доля гранулированного эндоплазматического ретикулума и увеличено количество свободных полирибосом (Бердинских, 1983), которые являются системой синтеза белков, необходимых для удовлетворения внутренних потребностей клетки. Не исключено, что к этому разобщению причастно рассмотренное выше снижение уровня CYP, поскольку есть, правда, давние сведения о возможном участии этого фермента во взаимодействии рибосом с эндоплазматической мембраной (Ohls-son, Jergil, 1977). Причиной преобладания свободных полирибосом в опухолевой клетке могут быть также уже рассмотренные в п. 2.3.4 деструктивные изменения элементов цитоскелета, в том числе микротрубочек. Эксперименты по изучению влияния колхицина, разрушающего эти структуры, на связь с мембраной полисом печени крыс показали: цитоплазматические микротрубочки необходимы для образования связанных с мембранами полирибосом. Действительно, после введения колхицина из фракции таких полирибосом исчезали мРНК, но не рРНК. Колхицин не влиял на содержание мРНК и рРНК во фракции свободных полисом (Walker, Whitfield, 1985).
Значительное преобладание в опухолевых клетках свободных полирибосом за счёт соответствующего уменьшения мембраносвязанных детерминирует ряд изменений, вносящих вклад в характерные черты «ракового» обмена.
Так, усиленный в гепатоме синтез ключевого фермента гликолиза гексокиназы происходит в основном на свободных полирибосомах (Нейфах и др., 1977). Это явление, связываемое авторами со сдвигами в энергетическом обмене малигнизированных клеток, несомненно, способствует усилению гликолиза. Синтез in vitro Cu-, Zn- и Mn-SOD печени крыс также осуществляется преимущественно в присутствии фракции свободных полирибосом (Hirano et al., 1985). Если это признать типичным явлением, то в опухолевых клетках содержание SOD должно быть увеличенным.С другой стороны, несмотря на некоторую противоречивость мнений, достоверно установлено, что активность этих ферментов в ряде типов неоплазм существенно ниже, чем в нормальных тканях (Marinaccio et al., 1990). Такое несоответствие между потенциально возможным повышением уровней SODs и низкой их активностью в опухолях может объясняться, в частности, данными об ингибировании указанных ферментов первичными продуктами ПОЛ мемб-ран (Гудзь и др., 1982), избыточное содержание которых постулируется нами в активно растущих участках опухоли. По-видимому, системы образования и детоксикации О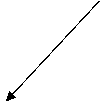 находятся в каком-то регуляторном взаимодействии: в норме активность SODs определяется уровнем О; при усилении же ПОЛ, сопровож-дающем пролиферацию клеток, разобщаются рибосомомембранные связи, т. е. создаются условия для появления свободных полирибосом и активизации синтеза на них SOD, призванных защитить клеточную среду от токсического действия О и его производных. Однако при канцерогенезе избыточное ПОЛ выходит за нормальные допустимые пределы изменения, и в этих условиях инактивации продуктами ПОЛ подвергаются и сами SODs.
находятся в каком-то регуляторном взаимодействии: в норме активность SODs определяется уровнем О; при усилении же ПОЛ, сопровож-дающем пролиферацию клеток, разобщаются рибосомомембранные связи, т. е. создаются условия для появления свободных полирибосом и активизации синтеза на них SOD, призванных защитить клеточную среду от токсического действия О и его производных. Однако при канцерогенезе избыточное ПОЛ выходит за нормальные допустимые пределы изменения, и в этих условиях инактивации продуктами ПОЛ подвергаются и сами SODs.
Другая часть негативных эффектов при дестабилизации микросомальных мембран вызвана дефицитом мембраносвязанных полирибосом. Наряду со снижением количества синтезируемых на них глутамат- и малатдегидрогеназы, цитохрома с и белков внешней мембраны митохондрий (Бердинских, 1983), о чём уже упоминалось в п. 2.1.7, в опухолевой клетке ещё более усугубляется ситуация с уменьшением содержания цитохромов CYP и b5, поскольку они синтезируются только на прочно связанных с мембранами полирибосомах (Apffel, 1982). Падает и синтез на них специфических для дифференцированных клеток белков. В качестве приспособительной меры синтез в опухолях некоторой части белков, предназначенных для секреции, «организуется» на свобод-ных полирибосомах. Однако такие белки почему-то оказываются «лишними» и подвергаются деградации (см. Берман, 1985).
Микросомальная гипотеза канцерогенеза (Apffel, 1982), постулируя необратимую дегрануляцию шероховатого эндоплазматического ретикулума канцерогенами и нарушение контроля над синтезом белка, связывает эти изменения с нарушением мембран микросом. Однако она не даёт ответа на главные вопросы: каким образом канцерогенные агенты, в частности химические, повреждают эти мембраны, каков конкретный механизм взаимодействия их с микросомами. С точки же зрения кислородно-перекисной модели, указанные изменения в мембранах микросом не первичны, не связаны прямо с канцерогенным воздействием, а есть лишь одно из следствий создающейся внутриклеточной гипер-оксии и последующей пероксидации липидов во всех мембранах.
Известная естественная функция микросом подвергать метаболическому превращению ксенобиотики может быть квалифицирована как соучастие микросом в кислородно-перекисном механизме канцерогенеза. Наиболее реально это в тех случаях, когда в ходе указанного превращения образуются АФК. К числу таких примеров можно отнести микросомную активацию тамоксифена и 4-гидрокситамоксифена в присутствии NADPH, сопровождающуюся генера-цией АФК. Последние приводят к образованию 8-OH-dG, с чем и связывается канцерогенное действие тамоксифена (Ye, Bodell, 1996). Действие гетероциклических аминов, одной из наиболее хорошо изученных групп канцерогенов, сопровождается образованием О в присутствии CYP или CYP-редуктазы и нередко окислительной модификацией яДНК (Sato et al., 1992; Маеда, Акаике, 1998). Если же при метаболическом превращении канцерогенов непрямого действия возникают конечные канцерогены, как, например, в реакции 2′-гидро-ксилирования никотина CYP2A6 и микросомами человека (Hecht et al., 2000), то объектом прямой атаки их, как отмечалось в п. 2.1.1, могут быть прежде всего митохондрии, а внутри них – дыхательные ферменты и мтДНК.
Считаем уместным указать здесь и на следующее возможное «биокибернетическое» обобщение. Как нам представляется, переходу нормальной клетки в состояние пролиферации способствуют обратимые премитотические гипероксия и усиление ПОЛ в мембранах эндоплазматического ретикулума, что приводит не просто к «разборке» этих мембран, но и к высвобождению связанных с ними рибосом. Феномен дестабилизации мембрано-полирибосомных комп-лексов может иметь общее принципиально важное регуляторное значение, связанное с реализацией триггерного принципа взаимосвязи размножения и дифференцировки клетки. Было постулировано, что триггерный принцип регуляции здесь организован по двухкаскадной схеме: на нижнем уровне так взаимодействуют отдельные транскриптоны генома, а на верхнем – целые подсистемы в составе глобальной транскриптоно-триггерной сети (Лю, Ефимов, 1978). Разобщение связи полирибосом с мембранами микросом в интенсивно размножающихся клетках рассматривается как механизм поддержания их пролиферации и одновременно ограничения в них возможностей для специализации. Реализация этого механизма, видимо, осуществляется просто путём регуляции соотношения свободных и связанных полирибосом в зависимости от интенсивности стимулирующих пролиферацию сигналов. Это соотношение, будучи показателем материальной базы на этапе трансляции и отражением фактического состояния во взаимодействии подсистемы управления клеточным циклом с подсистемами дифференцировки (специализации), может указывать на целесообразное в данный момент распределение рибосом для специального белкового синтеза и на нужды размножения и роста клетки.