Скорость передвижения (плавания) водных организмов
Таблица 36
| Животное | MjCCK |
| Барракуда (Sphyraena pictida). | 12,2 |
| Голубая акула (Carcharias glaucus) | 10,1 |
| Тунец (Thunnus thunnus) .................................... | 6.2 |
| Лосось (Saltno salar)..................................... | 6.0 |
| Форель (Saltno truita morplia furio) | 3,6 |
| Щука обыкновенная (Esox lucius) | 3.5 |
| Треска (Gadus morhua) ................................. | 2.5 |
| Плотва (Rutilus rutilus)................................. | 1.2 |
| Лещ (Abramis bratna)................................................................... | 0.9 |
| Налим (Lota lota).............................................. | 0.6 |
| Морской окунь (Scbastes rnarinus) | 0.4 |
Стандартная скорость плавания рыб принимается обычно как 4 Л. У мелких рыб при бросках эта скорость может намного превышать отношение к длине тела. Так как сопротивление движению пропорционально квадрату линейных размеров, а мощность (масса) мышц — кубу линейных размеров, крупные рыбы движутся при прочих равных условиях быстрее мелких.
Некоторые морские млекопитающие отличаются большой скоростью плавания. Так, для кита-сервала описана скорость 6 м/сек, для дельфинов до 10 м/сек и более.
Скорость передвижения в воде зависит от формы тела, характера движений туловища, хвоста и плавников, а также от гидродинамических особенностей тела рыбы, т.
е. от сопротивления, оказываемого водой при продвижении вперед. Между скоростью плавания рыбы и ее гидродинамическими свойствами существует значительный параллелизм (Шулейкин, 1966). Так, наименьшее сопротивление при движении оказывает вода телу дельфина, наибольшее медленно плавающей рыбе — морскому языку.Гидродинамические свойства плавающих организмов определяются, однако, не только формой тела, но особенностями его поверхности. Помимо гладкой поверхности, почти не создающей трения и вызывающей минимальное образование пограничного слоя движущейся вместе с плывущим телом воды, здесь большое значение имеет выделение слизи. Она выделяется специальными бокаловидными железами и имеет большое значение для увеличения коэффициента полезного действия плавающего животного. Мягкий демпферный слой подкожной жировой клетчатки у дельфинов также выполняет функцию гашения турбулентности.
Плавание осуществляется благодаря движениям тела, при которых поперечные колебания головы оказываются меньшими по сравнению с колебаниями хвоста. Большой движущей силой обладает хвост; он перемещается из стороны в сторону, проделывая синусоидный путь. Само тело рыбы в зависимости от вида в большей или меньшей степени проделывает синусоидные движения. Силы, действующие при сокращении туловища и движениях хвоста, изгибают хвост, но тем не менее хвост образует угол с движением тела по воде. Поэтому на хвост действуют и сила сокращения, «и упор о воду. Равнодействующая этих сил направлена вперед. Когда хвост перемещается влево, равнодействующая направлена вправо и наоборот, но средняя сила, измеренная на всем протяжении сокращения цикла, направлена точно вперед.
Мощность общей работы, выполняемой телом плывущей рыбы, является произведением скорости и силы сопротивления движению. Эта сила в свою очередь складывается из силы сопротивления воды движению сложно изгибающегося тела рыбы и силы внутреннего сопротивления изгибу тела. Кроме того, затрачивается энергия 362
на сопротивление распространяющейся вдоль тела волны.
Специальные исследования показали, что каждый сегмент тела рыбы обладает различной мощностью, и это соответствует и анатомической толщине расположенных на отдельном сегменте мышц (Шулейкин, 1968). Мышечный тяж, идущий вдоль тела рыбы, обладает в различных его участках (сегментах) разными биохимическими особенностями. Следовательно, плавание рыбы представляет исключительно интересный объект для изучения эволюции адаптаций мышечной системы к выполнению различных нагрузок.
Энергетический обмен у рыб при плавании был объектом многих работ отечественных и иностранных исследований. Поскольку многие рыбы существуют в проточной воде, плавание как способность противостоять течению является у них функцией непрерывной. Отсюда возникло понятие о так называемом активном обмене — обмене в толще движущейся воды (Ивлев, 1960). Эти величины активного обмена составляют около 200% от уровня обмена веществ в неподвижной воде.
Исследование мышечной деятельности (плавания) рыб производится в специальных гидродинамических камерах, где можно создавать различную скорость течения воды, поддерживать постоянную температуру и измерять содержание кислорода в воде (Fry a. Hart, 1948; Blazka a. oth., 1960; Brett, 1964), содержание углекислоты. Подобно исследованиям, проведенным на наземных организмах, главным образом на человеке, энергетическая оценка движения — плавания производится путем вычитания из величин газообмена, наблюдаемых во время плавания, величин так называемого стандартного обмена— более низких величин газообмена в состоянии полного покоя рыбы. Подобно фактам, хорошо изученным на человеке, потребление кислорода при плавании с достаточно большой скоростью не сопровождается полным удовлетворением кислородного долга, что установлено как при сопоставлении теоретически вычисленных величин потребления кислорода с наблюдаемыми фактически, так и при изучении величин кислородного долга (Brett, 1964; Smith, 1965). С возрастанием скорости газообмен повышается, однако имеется определенный энергетический оптимум скорости, когда газообмен, рассчи-
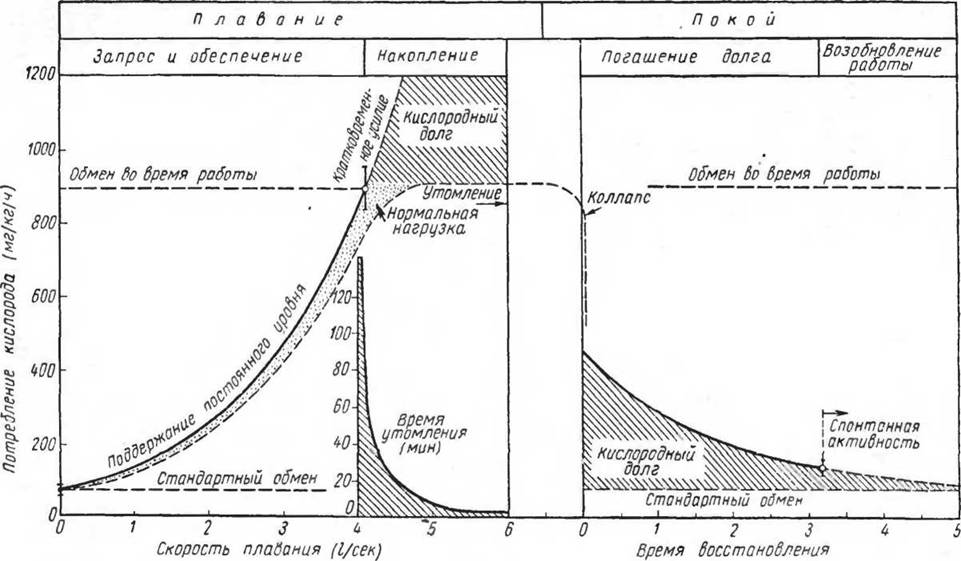
Рис.
79. Взаимоотношения между потреблением кислорода, скоростью движения (плавания), временем работы до утомления, кислородным долгом п восстановлением у красной нерки при температуре воды 15° С (по Brett, 1964)танный на пройденный путь, оказывается наименьшим. Этим величинам ряд исследователей придают большое значение, предполагая, что именно эти наиболее «экономные» скорости используются рыбами в процессе дальних миграций.
Одной из характерных черт мышечной энергетики рыб является очень длительная (после работ, проведенных до утомления) ликвидация кислородного долга, продолжающаяся более 3 ч. Нагрузка, при которой рыба может продолжать работу в устойчивом состоянии (steady state), не превышает величины 4 линейных ее размеров (4 Л) в секунду (Brett, 1964). На рисунке 79 приведены отношения потребления кислорода, скорости утомления и восстановления у нерки (Oncorhynchus nerka} весом 50 г.
Таблица 37
Потребление кислорода и скорость плавания при различных температурах среды у нерки (по Brett, 1964)
| Температура сречы (°С) | Вес тела | Потребление О5 (мг!к: ч) при работе | Потреблен не О2 (мг кг, ч) станлертный обмен | Критическая скорость плавайия | |
| г сек | 1'сек | ||||
| 5 | 37,6 | 514 | 41 | 1,76 | 3.26 |
| 10 | 32,9 | 627 | 60 | 1Л0 | 3,65 |
| 15 | 55,2 | 895 | 71 | 2,52 | 4,12 |
| 20 | 62,6 | 852 | 120 | 2,43 | 3,90 |
| 24 | 52,2 | 848 | 196 | 2,25 | 3,75 |
В последнее время для изучения энергетических затрат плавающих рыб стали применять косвенный метод расчета по содержанию жира в теле.
Эти пути исследования имеют большое значение при изучении миграций.Весьма интересные данные по плаванию рыб получены при различной температуре воды. Максимальная скорость плавания с повышением температуры воды возрастает. При этом стандартный метаболизм увеличивается с 41 до 196 мг1кг1ч\ расход же на плавание — с 514 до 948 мгікгіч, т. е. менее чем вдвое. Эти результаты получены после двухнедельной адаптации к температурам от 5 до 24° С и указывают иа значительную независимость коэффициента полезного действия скелетной мускулатуры рыбы от температуры среды и от предшествующих температурных воздействий, да и скорость плавания не возрастает с температурой выше 15° С.
Несмотря на большой интерес, проявляемый в последнее время к изучению морских млекопитающих, особенно дельфинов, какие-либо данные о физиологии плавания отсутствуют. Имеются только предположительные расчеты, исходя из наблюдений за скоростью и весом тела, свойствами поверхности тела.
Еще по теме Скорость передвижения (плавания) водных организмов:
- ОСОБЕННОСТИ ПЕРЕДВИЖЕНИЯ (ПРЕОДОЛЕНИЯ ПРОСТРАНСТВА) В ВОДНОЙ СРЕДЕ
- Скорость передвижения (плавания) водных организмов