ФИЗИОЛОГИЧЕСКИЕ ИЗМЕНЕНИЯ ПРИ ЗИМНЕЙ СПЯЧКЕ
Зимнее угнетение жизнедеятельности организма, носящее сезонный циклический характер, особенно ярко вы- ражено у млекопитающих, впадающих в зимнюю и летнюю спячку. Под спячкой понимают состояние резко пониженной жизнедеятельности животного, сопровождающееся изменением поведения: оно перестает двигаться и принимает ооычно характерную позу, соответствующую позе сна.
Наиболее ярким проявлением спячки является снижение температуры тела. Многие исследователи считают, что в основе физиологического механизма спячки лежит временное нарушение или выключение терморегуляции, так называемое состояние гетеротермии. Тщательное сопоставление уровня температуры тела у зимоспящих и незимоспящих млекопитающих по возможности близких в систематическом отношении видов обнаруживает у зимоспящих более низкий уровень температуры тела (см. табл. 4).Таблица 4
Температура тела у зимоспящих и незимоспящих млекопитающих
(в состоянии бодрствования)
| Зим» сняти? | t c | ІІеЗИМГіСІІЯІІІІіе | t c |
| Еж европейский (Eri- | около | Крот европейский | 39,4 |
| naceus curopaeus) . . . | 35,0 | (Talpa euro раса) . . . | |
| Сурок альпийский | 36,0 | Серая крыса-пасюк | |
| (Marrnola baibacina) . . | (Rattus norvegicus) . . | 37,6 | |
| Суслик крапчатый (Сі- | 36,5 | Заяц-русак (Lepus | 38,0 |
| iellus suslicus).................. | europeus) ......................... | ||
| Хомяк обыкновенный | 34,7 | Полевая мышь (Apode- | |
| (Cricetus cricetus) . . . | mus agrarius)................... | 38,5 | |
| Соня-полчок (Glis | 35,5 | Морская свинка (Pro- | |
| gtis).................................. | cavia cobaya) .................. | 38,6 | |
| Соня орешниковая | Собака (Cunis fainilia- | 38,5 | |
| (Muscardinus avcllanari- | ris).................................... | ||
| us).................................... | 35,7 | Шакал (Canis aureus) | 38,5 |
| Барсук (Meles meles) Медведь закавказский | 37,8 | Лисица (Vulpes vul- Pes).. | 39,0 |
| (Ursus arctos caucasicus) | 37,0 | ||
| Енотовидная собака | |||
| (Nyctereutes procyonoi- des)........................................ | 37,8 |
Многие исследователи указывают, что температура тела млекопитающих, впадающих в спячку, отличается большим непостоянством даже в период бодрствования,
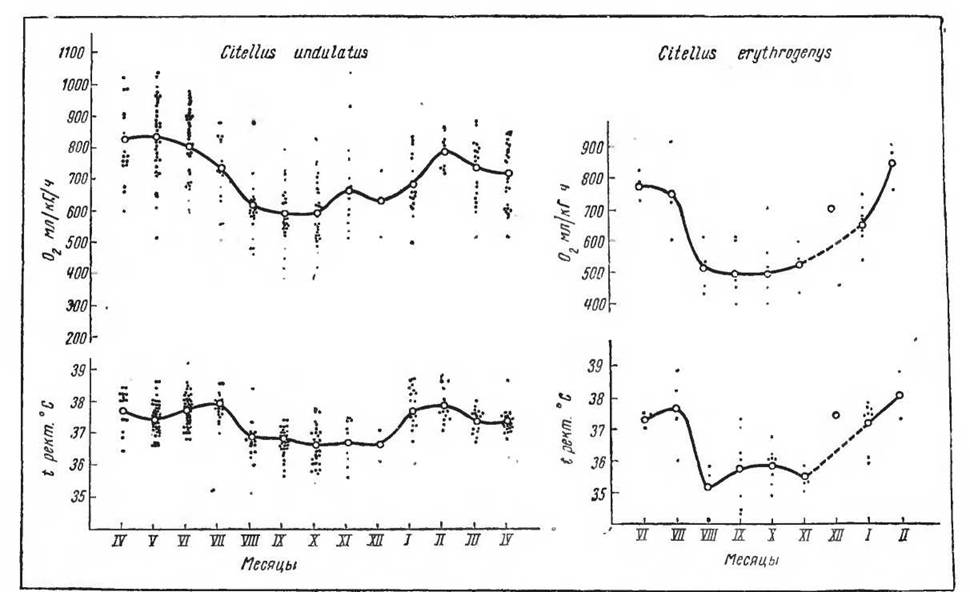
Рис.
32. Сезонные изменения температуры тела и потребления кислорода у двух видов сусликов' краснощекого (Citellus erythrogenys) и длиннохвостого (Citellus undulatus) (по Мнхневнч, 1969)например у ежен, сонь, сусликов. Гак, суточные колебания температуры тела ежа могут достигать около 2,5° С (Herter, 1933) , хомяка — от 2,1 до 7,0° С (Ilorwath, 1878; Kayser, 1939а, 19396, 1940а, 19406). Таким образом, у зимоспящих форм и в период бодрствования наблюдается более низкая температура тела, подверженная более значительным колебаниям под влиянием изменяющихся температур среды. У бодрствующих сусликов при круглогодичном содержании при комнатной температуре можно было наблюдать закономерные снижения уровня обмена веществ п температуры тела в месяцы, соответствующие периоду спячки в природе (рис. 32). У наземных хищных (см. табл. 4)—барехка, медведя, енотовидной собаки, для которых характерен лишь продолжительный зимний сон, температура тела также несколько снижена по сравнению с другими хищными млекопитающими (Слоним,
1952).
В период зимней спячки температура тела снижается, как правило, до уровня температуры окружающей среды, лишь на доли градуса превышая ее. Другим не меиее характерным проявлением спячки является резкое снижение обмена веществ (потребления кислорода, выделения углекислоты) в 10 и более раз по сравнению с состоянием бодрствования. Уровень обмена зависит от температуры среды, глубины спячки, но вообще он остается довольно постоянным па протяжении многих месяцев.
О величинах снижения теплопродукции во время зимней спячки у разных млекопитающих можно судить по таблице 5.
По-видимому, понижение обмена наступает в одинаковой мере у всех видов вне зависимости от величины поверхности тела. Следовательно, в состоянии зимней спячки закон поверхности тела теряет свое значение.
Наблюдается резкое замедление частоты дыхания, а иногда и прекращение его. В этом случае обмен газов осуществляется благодаря так называемым кардиопнев- матнческнм движениям, когда сердечные сокращения, также редкие, вызывают толчок, который способствует перемешиванию воздуха в воздухоносных путях, достаточный для того, чтобы удовлетворить резко сниженный кислородный запрос.
Так, например, у ежа частота дыхания в бодрствующем состоянии достигает 40—50 в минуту, а при впадении в спячку падает до 6—8 и даже доТеплопродукция у некоторых зимоспящих в летний (при бодрствовании) и зимний период (в спячке)
(по Kayser, из Eisentraut, 1956)
| Знмоспящие | Летом | Зимой | ||
| Вег тела G) | Калории (кг, -і) | Вес тел;: (г) | Калории (кг.ч) | |
| Сурок обыкновенный (Mar mot и bobac) . . . | 1868 | 2.1 | 2146 | 0,086 |
| Еж европейский (Eri- naceus curopaeus) . . . | 684 | 3.5 | 600 | 0.075 |
| Суслик малый (Citel- lus pygmaeus)................................ | 227 | 4.5 | 275 | 0,085 |
| Соня-полчок (Glis glis).......................................... | 127 | 5.0 | 130 | 0,069 |
| Соня орешниковая (Muscardinus avellanarius) | 19 | 13,0 | 23 | 0,170 |
1 в минуту (Suomalainen, 1944). У сурка частота дыхания понижается с 25 до 4—5 в 1 мин (Benedict a. Lee, 1938). Подобные же данные имеются для зимоспящих сусликов, летучих мышеи и т. д. Во время зимнего сна у хищных (барсука, медведя) частота дыхания заметно не снижается.
Тщательно изучены изменения дыхания орешниковой сони (Muscardinus avellanarius) при зимней спячке и пробуждении (Томилин, 1958).
Во время глубокой зимней спячки обнаружены дыхательные паузы продолжительностью до 11 мин. Такие паузы чередуются с сериями дыхательных движений. При пробуждении (повышении температуры тела) длительность дыхательных пауз возрастает, а число дыханий между паузами уменьшается; при понижении температуры имеют место обратные соотношения. При температуре среды около —5,5° С наступает пробуждение, и температура тела достигает 34,5— 36,0° С, т. е. температуры тела бодрствующего животного.Очень сильно изменяется во время зимней спячки кровообращение. Частота сердечных сокращений падает у ежа с 230 до 24 в минуту (Suomalainen a. Sarajas, 1951). У суслика частота пульса урежается в 8—10 раз
(Johnson, 1929). Минутный объем сердца может у суслика снижаться в 65 раз (Popovic, 1964). Резко замедляется во время спячки кровоток. Если время оборота крови у бодрствующего животного — кролика составляет 7— 8 сек, то у спящего сурка — 3—4 мин. Большое количество крови депонируется в селезенке. Кровяное давление падает с 70—72 мм Hg до 16 мм Hg в сонной артерии (Dubois, 1896).
Большой интерес представляют морфо-физиологические особенности сердца зимоспящих. Сердце зимоспящих
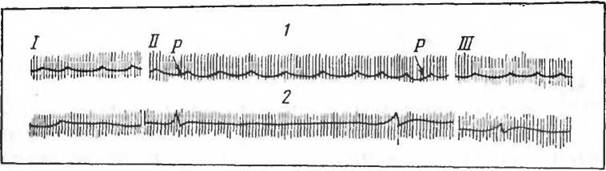
Рис. 33. Электрокардиограмма европейского ежа (по Suonialainen и Sarajas, 1951) I, II, III — отведения:
I — и состоянии бодрствования, 2 — в зимней спячке
млекопитающих способно к ритмическим сокращениям даже при 0° С, тогда как у незимоспящих его остановка происходит уже при температурах около 15° С (Lyman а. Chatfield, 1956; Sarajas, 1960; Johansson, 1967, 1968). При этом у зимоспящих никогда не наблюдается явлений фибрилляции желудочков при выходе из состояния гипотермии, что является обычным осложнением при попытках вывести незимоспящее животное из гипотермии, вызванной охлаждением. Эта очень интересная особенность сердца зимоспящих получила свое объяснение недавно. Оказалось, что клетки миокарда зимоспящих совершенно не иннервируются адренергическими волокнами.
Эта иннервация ограничивается лишь сосудами миокарда и повышение содержания катехоламинов, имеющее место при пробуждении, не вызывает явлений фибрилляции желудочков (Nielsen a. Owman, 1968). Во время спячки резко уменьшается количество мочи, а концентрация ее 4значительно возрастает.Терморегуляция и температурная чувствительность не полностью выключена. Резкое охлаждение зимоспяще/о животного обязательно вызывает пробуждение. /Можно думать, что в условиях зимней спячки и значительного выключения терморегуляционной активности сохраняется «сторожевой пункт» в центральной нервной системе, раздражение которого вызывает пробуждение.
Глубоким изменениям подвергается также качественная сторона обмена веществ. Так как удовлетворение пониженных энергетических потребностей организма во время спячки осуществляется за счет накопленных жиров, то в это время наиболее интенсивные изменения происходят в жировом обмене. В теле летучей мыши до зимней спячки было найдено 21,2 г жирных кислот на 100 г веса тела, а после 58 дней спячки — лишь 12,6 г (Reach, 1910).
Различные жиры, отложенные к концу лета, расходуются на протяжении зимней спячки неодинаково. Зимняя спячка сравнительно мало отражается на количестве внутреннего жира у тарбаганов; в этот период сильно расходуется подкожный жир. К концу спячки у летучей мыши (Myotis myotis) точка плавления жира оказалась повышенной (Mann, 1936). Изменяется также йодное число, кислотность жира и другие его химические и физические свойства.
Особую роль в протекании зимней спячки и в пробуждении играет скопление бурого жира, или бурой жировой ткани. До недавнего времени этой ткани приписывали роль своеобразной эндокринной железы. Однако в последние годы благодаря химическим и гистохимическим исследованиям роль бурого жира становится более ясной (Joel, 1964). Анатомически бурый жир располагается в области сердца, диафрагмы и в межлопаточной области вдоль позвоночника. Цвет бурой жировой ткани обусловлен скоплением сосудов, богатством митохондрий и цитохрома. В межлопаточной бурой жировой ткани суслика содержание цитохромов в 4 раза больше, чем в ткани мозга или мышц.
Бурый жир встречается не только у зимоспящих организмов, но у них он подвержен значительным сезонным колебаниям. Особенно значительно убывание бурого жира в процессе пробуждения от зимней спячки. Так, из общего содержания липидов в бурой жировой ткани сусли-
ка в процессе пробуждения исчезает около 45%, а по весу эта ткань теряет около 22%.
Потребление кислорода бурой жировой тканью очень значительно. У суслика (Citellus tridecimlineatus) оно здесь гораздо выше, чем в ткани почек того же животного (Hook a. Barron, 1941). Особенно ярко этот факт проявляется при инкубировании срезов бурой жировой ткани при разных температурах и при добавлении гормонов и ферментов (см. табл. 6).
Таблица G
Потребление кислорода срезами бурой жировой ткани и органов суслика при разных температурах и добавлении
гормонов и ферментов (по Joel, 1965)
| Тк.ІІІИ II орг; ЦЫ | Ии куб ’НИЯ при 37° С (.w.zO., 100 -иг свежей ткани час) | Инкубация при 8° С (% от скорости при 37° С) | Количество нар инкубации |
| Бурая жировая ткань без добавления гормонов ........ | 539 + 15 | 31+3 | 12 |
| + норадреналин . . . | 719+32 | 38+3 | 12 |
| + норадреналин + сукцинат ............................................. | 1260 + 00 | 23 + 5 | Г) |
| Белая жировая ткань без добавления гормонов ........ | 10 + 1 | 9 + 1 | 7 |
| + норадреналин . . . | 14+2 | ■ )±3 | 7 |
| Печень............................ | 228 + 18 | 20 ±1 | 4 |
| Диафрагма...................... | 214± 18 | 20 + 2 | 7 |
| Почка.............................. | 418 + 24 | 8.2 + 0,4 | .) |
Наблюдается резкое уменьшение содержания гликогена в бурой жировой ткани после пробуждения. Таким образом, бурая жировая ткань представляется в настоящее время субстратом повышенных окислительных процессов при пробуждении и источником повышенного теплообразования (подробно см. гл. V). Высказывается предположение, что в процессе разогревания используется бурый жир, непосредственно прилегающий к жизненно важным органам — сердцу, спинному мозгу. На ранних стадиях 145 разогревания (пробуждения) кровообращение оказывается особенно интенсивным в этих отделах бурой жировой ткани, что сопровождается и начальным повышением температуры. В последующие стадии разогревания бурая жировая ткань является, вероятно, источником значительного повышения содержания жировых кислот в крови.
Во время спячки дыхательный коэффициент падает до очень низких величин, что, однако, связано, по имеющимся данным, с удержанием некоторого количества углекислоты кальцием и магнием костей.
Существуют многочисленные доказательства значительных изменений углеводного обмена во время зимней спячки. Уже давно (Valentin, 1857; Bernard, 1859) установлено, что содержание гликогена в печени в период спячки увеличивается, а в период пробуждения падает. Содержание сахара в крови во время спячки резко снижается, доходя до 33 лш%, и повышается в период пробуждения до 180 мг% и более (Dubois, 1896); количество гликогена на протяжении спячки закономерно падает в печени, сердце, поперечнополосатой мускулатуре (Leduc et Lyman, 1952). Между уровнем сахара крови и температурой тела во время спячки наблюдаются прямые соотношения: чем ниже температура тела, тем больше выражена гипогликемия (Endrews, 1930; Фердманн и Файи- шмидт, 1932; Suomalainen, 1944). При пробуждении уровень молочной кислоты в крови нарастает (Фердманн и Файншмидт). Таким образом, для зимней спячки характерно своего рода блокирование углеводных резервов в организме. С пробуждением начинается их интенсивное расходование. Этому способствует энергичная мышечная деятельность — дрожание, связанное с пробуждением. Что касается обмена азота в организме зимоспящего, то в период спячки он резко снижается.
Своеобразным глубоким изменениям во время зимней спячки подвергаются процессы регуляции постоянства внутренней среды (гомеостазиса).
Еще в конце XIX столетия было установлено (Dubois, 1898), что в крови спящих животных (сурков) обнаруживается повышенное содержание углекислоты. Это дало повод этому исследователю рассматривать спячку (да и сон вообще), как проявление наркоза, вызванного углекислотой (карбонаркоза). Резкое нарастание содержания углекислоты связано с понижением возбудимости дыхательного центра. В последние годы установлен ф.акт значительного повышения щелочных резервов крови у спящего ежа (Филатова, 1946). В поддержании желочно- кислотного равновесия в период зимней спячки играет важную роль и выделение значительного количества кислого желудочного сока. Благодаря этому освобождается значительное количество щелочных оснований крови, связывается дополнительное количество угольной кислоты и уменьшается возбудимость дыхательного центра. По-видимому, в основе резкого сокращения частоты дыхания и уменьшения легочной вентиляции во время спячки лежат оба механизма — понижение возбудимости дыхательного центра и увеличение щелочных резервов крови (Персон, 1950). Изменяются и физико-химические свойства крови— удлиняется время свертывания. Механизм происхождения этих явлений пока не выяснен.
Изменения в нервной системе и соответственно рефлекторной деятельности имеют в основе выключение высших отделов центральной нервной системы: коры, подкорковых узлов, области промежуточного мозга. Все же сложные координации физиологических процессов, связанные с деятельностью продолговатого мозга, сохраняются. Больше того, во время спячки проявляются ярко выраженные рефлексы, которые отсутствуют в бодрствующем состоянии животного. Все эти рефлексы относятся к поддержанию позы в состоянии спячки. Обычно спящее животное принимает характерную позу: ежи сворачиваются в клубок благодаря сокращению специальной мускулатуры; для летучих мышей характерно подтягивание иа задних конечностях (во время спячкн они висят вниз головой). Установлена точная локализация этих сложных рефлексов в области продолговатого мозга. Таким образом, можно считать, что глубокая спячка характеризуется отнюдь не выключением, а усилением деятельности отдельных участков продолговатого мозга (Mersbacher, 1903).
У многих млекопитающих во время спячки сохраняется способность производить некоторые движения. Особенно ярко это выражено у летучих мышей. Если висящую головой вниз в состоянии глубокой спячки летучую мышь поместить в клеточку, а затем перевернуть ее таким образом, что животное окажется головой вверх, то можно на- блюдагь медленное переползание без одновременного повышения температуры тела и обмена, т. е. без пробуждения животного. Многие исследователи обращают внимание па перелеты летучих мышей с места па место в пещерах во время зимней спячки. По-видимому, здесь имеют место самые различные степени пробуждения животных. Однако, несмотря на значительные передвижения внутри зимних убежищ, летучие мыши во время спячки пищи не принимают. Вопрос этот требует дополнительных исследований и представляет большой физиологический интерес для изучения уровней двигательных координаций.
Относительно высшей нервной деятельности во время зимней спячки сказать что-либо трудно. Опыты с попытками выработать условные рефлексы во время зимней спячки не проводились. Ранее же образованные условные рефлексы во время спячки выпадают.
Имеются данные о том, что результаты дрессировки животных исчезают даже после кратковременной зимней спячки. Так, хомяки, приученные брать пищу из рук человека, теряли эту способность даже после короткой спячки и к ним возвращалась их природная дикость (Portig, 1950).
У ежа после кратковременной спячки наблюдалось исчезновение искусственного двигательного условного рефлекса па время кормления (в утренние часы) (Филатова, 19496). У летучей мыши (ночница, Nyctalus noctula) хорошо выработанный двигательный условный рефлекс на место кормления исчез после зимней спячки, продолжавшейся только одни сутки со снижением температуры тела животного до 3,5° С (Эйзеитраут, 1956). Исчезновение всех явлений приручения после кратковременной зимней спячки наблюдал этот же автор и у ушастого ежа. В этом отношении зимняя спячка сильно отличается от влияний гипотермии, после которой (Калихман, 1949; Malmejac et al., 1956) глубоких изменений условнорефлекторной деятельности не обнаруживается. Механизм такого рода явлений заслуживает самого тщательного изучения.
Однако во время зимней спячки у летучих мышей, по крайней мере, сохраняется суточный (циркадный) ритм обмена веществ. Он проявляется только в определенных температурных условиях, но отражает режим покоя и бодрствования, имевший место в летний период (Попугаева и Слоним, 1953).
Следовательно, можно думать, что образованные условные рефлексы на время частично сохраняются и во время самой зимней спячкн, по исчезают при пробуждении. Но, если предположить, что кора головного мозга во время спячкн выключена, то замыкание этой связи, очевидно, осуществляется за счет нижележащих центров го ловного мозга.
Вопрос о возможности сохранения памяти во время зимней спячкн в последние годы вновь привлек внимание исследователей в связи с электрофизиологическими данными о выпадении во время зимней спячки электрической активности мозга, лежащей в основе циклического возбуждения— реверберации. Реверберация, по мнению многих исследователей, является механизмом удержания кратковременной, образной памяти. В опытах над европейскими сусликами (Citellus citellus), у которых осенью были образованы навыки прохождения через лабиринт, было показано, что эти навыки хорошо сохраняются после длительной зимней спячкн на 32 и день после пробуждения. Это приводит к общему выводу, что долговременная память связана со структурными изменениями в нервной системе и сопровождается возрастанием содержания РН1\ и белковых субстанций в мозг) (Mihailovic, Petrovic-Miriic, Protic a. Divac, 1968). Значительное повышение содержания РШ\ в гиппокампе было обнаружено при пробуждении из спячки у краснощекого суслика (Citellus erythrogenys)-, после полного пробуждения содержание РНК не отличалось от содержания во время спячки (Семешина, 1969). Таким образом, различные формы памяти в разной мере сохраняются как при искусственной гипотермии, так и при естественной зимней спячке.