Гипотезы о природе угнетения условных рефлексов под влиянием необычных раздражений.
Так как временнь е связи условных рефлексов находятся в коре большого мозга, можно было предположить, что условные рефлексы угнетаются под влиянием необычных раздражений путем торможения временных связей в коре большого мозга.
К такому именно заключению пришел Павлов (15). Он назвал это торможение внешним и предположил, что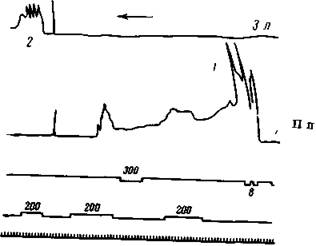 |
Рис. 167. Влияние электрического раздражения правой передней ноги на условный рефлекс задней левой ноги на тон 200 гц
В опыте 1 сначала производится электрическое раздражение, а затем на фоне последействия дважды условный тон 200 гц и один раз дифференцированный тон 300 гц верхний сигнал с обозначением 300) Условный звук усиливает оба раза это последействие, а своего рефлекса на задней левой ноге не дает, необычный же тон не действует В опыте г условный тон дает нормальный эффект на задней левой ноге
когда какой-либо пункт коры приходит в деятельное состояние под влиянием внешних или внутренних раздражений, то это сейчас же ведет к понижению возбудимости или к полному исчезновению ее в других пунктах благодаря перетягиванию возбуждения из этих пунктов. Впоследствии Павлов объяснял явление внешнего торможения отрицательной индукцией из первично возбужденного очага, т. е. предполагалось, что очаг возбуждения в коре большого мозга сопряженно производит торможение всех других очагов возбуждения коры. Таким образом, по Павлову, все тормозящее действие необычного раздражения протекает в коре большого мозга (Павлов, 15).
Долгое время мы также придерживались мнения, что угнетение условных рефлексов происходит путем торможения корковой части рефлекторной дуги (Беритов, 1927—1932). Позднее мы даже находили подтверждение этому положению в физиологических открытиях. Именно, когда было установлено, что в коре большого мозга также происходит общее торможение, обусловленное медленными потенциалами дендритного происхождения, нами было предположено, что отрицательное действие необычного раздражения на условный рефлекс вызвано главным образом общим торможением коры.
Предполагалось, что любое действие внешнего мира производит возбуждение определенных нервных кругов коры и одновременно, под влиянием коллатералей этих нервных кругов, производит активирование апикальных и базальных дендритов соседних пирамидных нейронов, которое приводит к общему торможению остальной коры (Беритов, 16, 17).Представление о корковом происхождении торможения условного рефлекса при более или менее интенсивных необычных раздражениях не может быть полностью оправдано. Мы знаем, что при подобных раздражениях происходит общее торможение рефлекторной деятельности почти по всей центральной нервной системе. Это торможение захватывает ядра как головного мозга, так и спинного. Например, у щенков при сдавлении кожи на голове
385
происходит небольшое движение головы и общее торможение всей мускулатуры, с угнетением как фазных, так и тонических рефлексов. То же происходит при акте еды, при вытягивании и сжимании корня хвоста и вообще при любых сдавлениях кожи. Если эти раздражения вызывают определенное
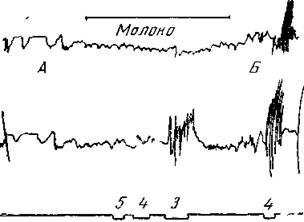
Рис. 168. Влияние еды на оборонительный рефлекс от электрического раздражения левой передней лапы
Щенок без новой коры большого мозга на 8-й День после операции Щенок подвешен на полотенце Лапы вытянуты вниз через отверстия в полотенце Верхняя запись — правая передняя лапа; средняя — левая передняя, нижняя — отметка раздражения левой передней лапы (цифры обозначают расстояние между индукционными катушками, см) В опыте А — раздражение лапы при силе тока в 5 и 4 см во время лакания молока не вызывает заметной реакции, а при 3 си производит небольшую реакцию только на раздражаемой лапе. В опыте Б, когда собака не ест, раздражение 4 см дает хорошо выраженную общую реакцию с сильным движением как раздражаемой, так и другой лапы
рефлекторное движение, то они одновременно производят угнетение других рефлексов, а также дыхания, голосовой реакции и «спонтанных» вздрагиваний.
Характерно, что это одинаково наблюдается как на нормальных животных, так и на животных, лишенных новой коры (Беритов, 6).Так, общее угнетение прирожденных поведенческих реакций было показано на щенках без новой коры во время акта еды (рис.168) при сдавлении кожи головы (рис. 169).
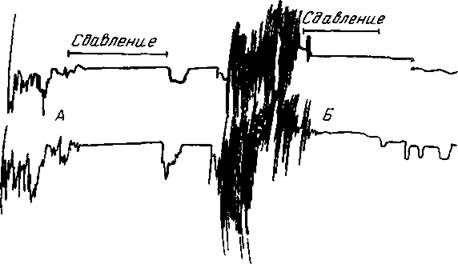
Рис. 169. То же животное
Обозначения те же, что на рис 168 А—во время общих освоСодительных движений и визга сдавливается кожа на голове, тотчас же прекращаю ся движения и визг, Ь - ко время «спонтанных» фазных движений по типу бега то же сдавленно прекращает его, при этом собака перестает визжать
386
Итак, факты свидетельствуют, что торможение условных рефлексов под влиянием необычных раздражений у нормальных животных может происходить в полной мере без участия коры большого мозга посредством общего торможения в спинном мозге и в стволе головного мозга. Так как общее торможение в стволе головного мозга должно осуществляться через ретикулярную формацию ствола мозга, в спинном мозге — через желатинозную субстанцию, надо прийти к заключению, что и наблюдаемое у нормальных животных торможение условных рефлексов должно происходить главным образом посредством этих образований.
Еще по теме Гипотезы о природе угнетения условных рефлексов под влиянием необычных раздражений.:
- Гипотезы о происхождении тормозящего действия условного раздражения.
- Роль ретикулярной формации в угнетении условных рефлексов.
- Роль желатинозной субстанции в угнетении условных рефлексов.
- О тормозящем последействии посторонних раздражений на условный рефлекс.
- Образование условных рефлексов на сложное раздражение.
- Характеристика условного рефлекса на комплекс раздражений.
- Образование условного рефлекса путем обратного сочетания индифферентного раздражения с безусловным.
- Происхождение условного рефлекса на комплекс раздражении.
- 6. Условные рефлексы на комплекс раздражений
- Влияние сознательной деятельности на условные и безусловные оборонительные рефлексы.
- Угашение условного рефлекса вследствие длительного неприменения условного раздражителя.
- Скорость образования условного рефлекса в зависимости от местонахождения условного раздражителя.
- Рецепторная дифференциация условных рефлексов.
- Генерализация условных рефлексов.
- О взаимодействии однородных условных рефлексов.
- Вестибулярные условные рефлексы.
- Генерализация условного рефлекса.
- Образование условных рефлексов высшего порядка.
- О взаимодействии разнородных условных рефлексов.
- Восстановление условных рефлексов.