ФИЗИОЛОГИЧЕСКИЕ АДАПТАЦИИ ПРИ мышечной деятельности
Поскольку всякая мышечная деятельность животного сопровождается увеличенным энергетическим расходом, наиболее общей чертой является здесь адаптация физиологических систем, направленных на обеспечение организма кислородом.
Поэтому наибольшие сдвиги в процессе адаптации происходят в функции дыхания, кровообращения и в дыхательной функции крови.Между величиной выполняемой животным работы как в короткие промежутки времени, так и на протяжении, например суток, и общим энергетическим расходом существуют прямые соотношения. Однако только для очень немногих форм деятельности известны энергетические характеристики; большинство сложных актов поведения и длительной работы животных с этой стороны еще совершенно не изучены.
Мышечная деятельность по своей интенсивности и общей величине выполненной работы распределяется в течение жизни животного совершенно неравномерно. Особенно интенсивна мышечная деятельность в определенные периоды жизни и в определенные сезоны. Так, большая мышечная работа производится животными при кочевках, миграциях и перелетах. С другой стороны, например, период беременности у самок многих грызунов, сопровождается резким ограничением мышечной активности. Большую мышечную работу производят животные при постройке нор и укрытий, а также в процессе запасания корма. Наконец, резкие возрастные отличия, связанные с повышенной игровой деятельностью молодняка, характеризуют большинство млекопитающих.
Мышечная деятельность по своей интенсивности и суммарному эффекту (в кг)м работы) величина постоянная для отдельных видов. Особенно это характерно для многих хищных, которые даже в условиях клеточного содержания в зоопарках выполняют большую мышечную работу, ритмически передвигаясь по клетке в течение многих часов в сутки (многие куньи, рыси, гиены, ЛЬВЫ и т. д.). Физиологический механизм, лежащий в основе этой «произвольной» деятельности, совершенно не ясен.
Во всяком случае, он никак не связан с пищедобывательной, оборонительной или какой-либо другой формой деятельности. Происхождение этой деятельности также неясно, как и происхождение игровой деятельности молодого животного.Физиологические изменения, происходящие при мышечной работе, а равно и приспособление к выполнению
мышечной работы различной интенсивности, скорости и продолжительности изучены главным образом на человеке в связи с проблемами физиологии труда и спорта. При трактовке многих фактов, касающихся мышечной деятельности животных, как домашних, так и диких, приходится опираться на этот большой и хорошо систематизированный материал (Atzler, 1927; Бейнбридж, 1927; Кекчеев, 1931; Конради, Слоним, Фарфель, 1934; Виноградов, 1941; Крестовников, 1951; Зимкин, 1956; Виноградов, 1958).
При мышечной работе наблюдается увеличение содержания эритроцитов и НЬ в крови. Это обычно связывается с сокращением селезенки и выбрасыванием крови из кровяных депо в периферическое кровяное русло. Объем такой депонированной крови может содержать до 40% всей крови. По-видимому, у разных млекопитающих роль отдельных депо различна. У человека это в основном кожа, у собаки — селезенка и сосуды печени. Сдвиг в содержании эритроцитов объясняется тем, что депонированная кровь содержит больше эритроцитов, чем циркулирующая. Кроме того, усиливается и новообразование крови, что выражается в возрастании числа ретикулоцптов.
Наиболее ярки и доступны для наблюдения изменения частоты пульса. При мышечной работе частота пульса возрастает параллельно ее интенсивности, т. е. параллельно потреблению кислорода. При очень интенсивной мышечной работе последнее происходит ие только во время выполнения работы, но и после ее окончания. Соответственно этому и пульс, учащенный во время работы, приходит к исходным величинам спустя несколько минут (иногда 50—60) после окончания работы. Установлено, что у тренированных людей и животных пульс учащается меньше, чем у нетренированных.
У тренированного человека при мышечной работе резко увеличивается объем каждой систолы—ударный объем сердца, а частота пульса сохраняется на более низком уровне.Факт уреженного пульса у тренированного человека, объясняемый обычно повышением тонуса блуждающего нерва, можно наблюдать и у животных, выполняющих очень интенсивную мышечную работу и хорошо приспособленных к ней. Так, частота пульса у зайца составляет около 70 в 1 мин, у кролика, не выполняющего 376
столь интенсивной работы, как заяц, частота пульса в покое — около 180 в 1 мин (Аршавский, 1958). Такие же соотношения были обнаружены при сравнении частоты пульса каракульских и романовских овец и гибридов 1 генерации архара и каракульской овцы. Частота пульса у каракульских овец была около 100, а у гибрида в покое — около 70. Это находит свое отражение и при сравнении лошадей различных пород. Так, частота пульса у шотландского пони весом 284 кг около 42,9 в 1 мин, а у першерона весом 646 кг — также около 42,8 (Brody, 1949). Таким образом, несмотря на разницу в весе, приспособленный к интенсивной работе пони имеет относительно замедленный пульс. Частота пульса у очень крупных животных невелика — у слона она равна 31 в 1 мин (Brody, 1945).
Кровяное давление во время мышечной работы возрастает. Это касается прежде всего систолического давления, возрастание которого находится в прямой зависимости от интенсивности мышечной работы. Диастолическое кровяное давление падает или поддерживается на постоянном уровне. Разность между величинами систолического и диастолического давления — пульсовое кровяное давление, как правило, возрастает. Существует определенная зависимость между величиной пульсового кровяного давления и минутным объемом сердца.
Изменения дыхания при мышечной деятельности сопровождаются увеличением частоты и глубины дыхания, что позволяет увеличить минутный объем дыхания или легочной вентиляции. Как правило, учащение дыхания при мышечной работе сопровождается углублением его; однако есть основания считать, что при тренировке и в дыхании наблюдаются такие же изменения, как и в минутном и ударном объеме крови.
Частота и глубина дыхания зависят от интенсивности выполняемой работы. У тренированных людей увеличиваются и частота и глубина, у нетренированных — главным образом частота дыхания.Частота дыхания у животных, приспособленных к интенсивной мышечной деятельности, также несколько ниже, чем у домашних, мало приспособленных. Так, частота дыхания у гибридов архарокаракуль оказывается более редкой, а объем выдоха большим, чем у овец каракульской или романовской породы.
Замечательным свойством дыхательной деятельности и сердечнососудистой системы тренированного организма является способность к поддержанию содержания кислорода в артериальной крови на постоянном уровне даже при выполнении довольно интенсивной (но, конечно, не предельной, истощающей) мышечной работы. Измеренное с помощью осигемометра насыщение артериальной крови у тренированного человека показывает почти постоянный уровень как при интенсивной, так и при умеренной работе. У нетренированного человека в первые минуты после начала работы насыщение кислородом артериальной крови значительно падает (Войтке- вич, 1952). К сожалению, проверка этих данных в приложении к диким животным, способным к выполнению интенсивной мышечной деятельности, не производилась.
Поддержание постоянства внутренней среды и прежде всего постоянства состава крови и реакции осуществляется за счет выведения продуктов мышечного обмена (молочной кислоты, СОг) из организма почками и потом, а СО2 — легкими. Однако огромное значение для поддержания pH крови имеют буферные системы крови, которые лучше всего могут быть изучены суммарно как щелочные резервы крови. Щелочные резервы крови (способность крови связывать угольную кислоту) у человека и животных разной тренированности к мышечной работе также различны. У тренированных они повышены и сохраняют этот высокий уровень и при выполнении мышечной работы (Конради, Слоним, Фарфель, 1935). Повышенная резервная щелочность встречается у животных, выполняющих интенсивную мышечную работу. Северный олень обладает большой кислородной емкостью и более высокими щелочными резервами.
Кривая диссоциации оксигемоглобина сдвинута вправо, что обеспечивает интенсивный перенос кислорода и отдачу его тканям (Крепе, 1936; Антелидзе и Барбашова, 1938). Количество крови у северного оленя превышает наблюдающееся у домашних животных (от 11 до 14% к весу тела), тогда как у других сельскохозяйственных животных эти отношения не превышают 10% (Городецкий, 1959). Высокие щелочные резервы крови наблюдаются также у таких животных, как лошадь — 72%, осел — 66%, собака — 56% и, наконец, у человека — около 60%.Низкие щелочные резервы характерны для животных с малой активностью: кролик — 48%, кошка — 46%, мышь белая — 45%, морская свинка — 36% (Меньшикова и Пигарева, 1945). Любопытно, что низким щелочным резервом обладает романовская овца (по данным тех же авторов, около 45%). Это можно объяснить малой мышечной активностью овец этой породы, разводимых почти исключительно в стойловых условиях.
Содержание угольной ангидразы — фермента, расщепляющего водный раствор угольной кислоты, находится в обратной зависимости от величин щелочных резервов крови.
Существуют многочисленные исследования содержания сахара в крови при мышечной деятельности (Конради, Слоним, Фарфель, 1935; Виноградов, 1941 —1958). В качестве общей схемы, одинаково приемлемой для человека и животных, можно принять следующие положения. При коротких, хотя и интенсивных мышечных нагрузках, наблюдается возрастание содержания сахара в крови, особенно ярко выраженное в начале работы. Эю явление отражает картину мобилизации гликогена, расходуемого в первую очередь как энергетический материал для мышечной деятельности. При более продолжительной (хотя и не предельной по напряжению) мышечной деятельности наблюдается снижение содержания сахара в крови. Это явление отражает, вероятно, процессы начинающегося истощения запасов гликогена в организме. Оно связано также и с ресинтезом углеводов из молочной кислоты, происходящим на протяжении продолжительной и не слишком тяжелой мышечной работы. По-видимому, при более или менее продолжительной работе можно наблюдать как начальную фазу повышения содержания сахара в крови, так и фазу понижения, связанную с нарастающим истощением организма.
Эти факты, полученные на человеке, нашли свое подтверждение и при исследованиях, проведенных на лошадях как в процессе коротких беговых испытаний, так и при длительных переходах.Мышечная деятельность — полет, сопровождается у летучих мышей разогреванием, повышением температуры тела и возрастанием содержания сахара в крови. Во время длительного полета содержание сахара в крови падает.
По интенсивности работы и по возможностям обеспечения ее выполнения доставкой кислорода все мышечные работы могут быть разделены на 2 группы: истощающая работа, протекающая с нарастанием кислородного долга и требующая его устранения после окончания работы (или перехода на более легкую работу, например с бега на шаг), и работа, когда количество продуктов мышечного метаболизма (молочной кислоты), образующееся б единицу времени, не превышает того количества, которое может быть устранено окислением или восстановлением в исходные вещества (гликоген). Состояние такого равновесия получило название устойчивого состояния (steady state). Кроме величины выполняемой работы, оно зависит еще от мощности дыхательной и циркуляторной систем организма.
Однако такое состояние наблюдается лишь при сравнительно нетяжелых, а главное не очень интенсивных мышечных работах. При очень тяжелых, истощающих работах потребность работающих мышц в кислороде превышает возможности доставки его с помощью аппаратов дыхания и кровообращения. В этих условиях в результате несоответствия между кислородным запросом тканей и его доставкой образуется кислородный долг. Нарастание этого долга на протяжении самой работы ограничивает ее продолжительность, а кислородный долг ликвидируется после работы в результате усиленного дыхания и восстановительных процессов в мышцах.
Выше уже указывалось, что по особенностям выполнения мышечной работы организм можно характеризовать по силе, быстроте (скорости) и выносливости. Каждое из этих качеств в различной степени представлено в отдельных мышечных спортивных упражнениях, каждое из них в различной степени представлено у животных с разной экологической специализацией.
Картины кривых диссоциации оксигемоглобина у лошади и верблюда различны (Крепе, 1936). Кривая диссоциации лошади характеризовалась значительной крутизной падения, что в ее разрядной части (при рО2 40) обеспечивает быструю отдачу кислорода оксигемоглобином в тканевых капиллярах. Такой тип кривой диссоциации обеспечивает обильное снабжение тканей кислородом при резко возросшем кислородном запросе. Тип кривой диссоциации у верблюда оказался более
пологим, с меньшей возможностью отдачи кислорода тканями и с большей способностью к фиксации его кровью. Первый тип кривой обеспечивает быстроту и большую интенсивность мышечной деятельности, второй— медленную устойчивую работоспособность на длительное время (рис. 82).
Характер мышечной деятельности и работоспособность животного в значительной мере определяются также и морфологическими особенностями скелетной мускулатуры. В организме животного имеются мышцы двух
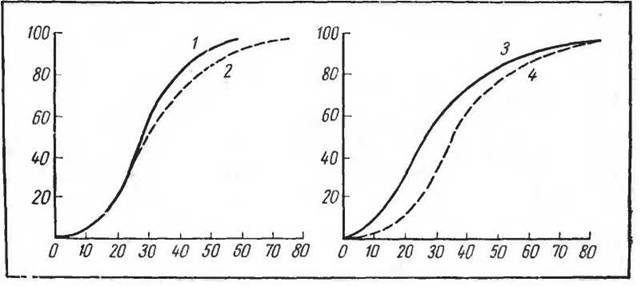
Рис. 82. Диссоциация оксигемоглобина у животных с разными типами мышечной деятельности (по Крепсу, 1936). По оси ординат — содержание оксигемоглобина (%); по оси абсцисс — напряжение кислорода (мм II):
/—лама. 2— верблюд, 3 — лошадь, 4 — северный олень
родов: мышцы белые, главным образом быстро сокращающиеся, и мышцы красные — тонические, обладающие свойствами более медленного сокращения и более развитыми свойствами поддержания мышечного напряжения— тонуса. Последнее особенно важно при поддержании позы и характеризуется малым энергетическим расходом, малой утомляемостью и малой интенсивностью центральной импульсации (Виноградов, 1958).
У лошадей количество желудочного сока, отделяемого во время мышечной работы, уменьшается, но после окончания работы быстро восстанавливается (Курмлов, 1955). Практически это находит свое отражение в том, что при мышечной работе интенсивностью около
2 .млн. кг/м в сутки наблюдается снижение переварива- емости сено-овсяных рационов. Последнее по клетчатке составляло 9%, а по жиру 7% (Чалюк, 1950). Однако это положение может быть правильным только для тех организмов, у которых процесс еды происходит или по окончании пищедобывательной деятельности (например, у хищника после охоты) или у животных, питающихся на одном месте после поисков корма (например, питающихся падалью, всеядных организмов).
Движения обезьян, направленные на добывание пищи (движения передних конечностей), всегда сопровождаются отделением слюны (Уголев, Волкова, Корневищ 1953). У лисицы при раскапывании норы грызунов наблюдалось увеличенное слюноотделение (Уждавини).
Мышечная деятельность оказывает очень большое влияние на терморегуляцию организма. Поскольку мышечные сокращения связаны со значительным теплообразованием и при выполнении работы в условиях средних температур (ие говоря уже о высоких), всегда имеется положительный баланс тепла. Это находит свое отражение в том, что температура тела, как правило, при интенсивной мышечной деятельности повышается. У человека такое повышение температуры тела может наблюдаться при работе значительной интенсивности, у животных — в зависимости от интенсивности работы и видовых особенностей животного. Наибольших сдвигов температура тела во время работы достигает у летучих мышей, являющихся практически пойкилотермными организмами в периоды отсутствия работы (полета).
Очень сильно повышается температура тела при мышечной работе у низших обезьян. Повышение температуры тела после бега в третбане может достигать 2,5— 3°С. Особенно ярко это выражено у молодых животных— подростков, отличающихся большой подвижностью.